 Бизнес-Блог Блоготей Бизнес-Блог «Блоготей» — информационный блог о бизнесе и финансах. Полезные статьи и новости из мира бизнеса, финансов, банков, экономики, инвестиций.
Бизнес-Блог Блоготей Бизнес-Блог «Блоготей» — информационный блог о бизнесе и финансах. Полезные статьи и новости из мира бизнеса, финансов, банков, экономики, инвестиций. 
Выписка по ИНН/ОГРН с печатью ФНС онлайн за 1 минуту
Политика конфиденциальности (далее – Политика) разработана в соответствии с Федеральным законом от 27.07.2006. №152-ФЗ «О персональных данных» (далее – ФЗ-152). Настоящая Политика определяет порядок обработки персональных данных и меры по обеспечению безопасности персональных данных в сервисе vipiska-nalog.com(далее – Оператор) с целью защиты прав и свобод человека и гражданина при обработке его персональных данных, в том числе защиты прав на неприкосновенность частной жизни, личную и семейную тайну. В соответствии с законом, сервис vipiska-nalog.com несет информационный характер и не обязывает посетителя к платежам и прочим действиям без его согласия. Сбор данных необходим исключительно для связи с посетителем по его желанию и информировании об услугах сервиса vipiska-nalog.com.
Основные положения нашей политики конфиденциальности могут быть сформулированы следующим образом:
Мы не передаем Ваши персональную информацию третьим лицам.
Собираемая информация
Мы собираем персональную информацию, которую Вы сознательно согласились раскрыть нам, для получения подробной информации об услугах компании. Персональная информация попадает к нам путем заполнения анкеты на сайте vipiska-nalog.com. Для того, чтобы получить подробную информацию об услугах, стоимости и видах оплат, Вам необходимо предоставить нам свой адрес электронной почты, имя (настоящее или вымышленное) и номер телефона. Эта информация предоставляется Вами добровольно и ее достоверность мы никак не проверяем.
Использование полученной информации
Информация, предоставляемая Вами при заполнении анкеты, обрабатывается только в момент запроса и не сохраняется. Мы используем эту информацию только для отправки Вам той информации, на которую Вы подписывались.
Предоставление информации третьим лицам
Мы очень серьезно относимся к защите Вашей частной жизни. Мы никогда не предоставим Вашу личную информацию третьим лицам, кроме случаев, когда это прямо может требовать Российское законодательство (например, по запросу суда). Вся контактная информация, которую Вы нам предоставляете, раскрывается только с Вашего разрешения. Адреса электронной почты никогда не публикуются на Сайте и используются нами только для связи с Вами.
Мы никогда не предоставим Вашу личную информацию третьим лицам, кроме случаев, когда это прямо может требовать Российское законодательство (например, по запросу суда). Вся контактная информация, которую Вы нам предоставляете, раскрывается только с Вашего разрешения. Адреса электронной почты никогда не публикуются на Сайте и используются нами только для связи с Вами.
Защита данных
Администрация Сайта осуществляет защиту информации, предоставленной пользователями, и использует ее только в соответствии с принятой Политикой конфиденциальности на Сайте.
ФНС > Запросы > Подготовка заявления на выписку из ЕГРЮЛ/ЕГРИП
В программе «СТЭК — ТРАСТ» предусмотрена подготовка заявления на выписку из Единого государственного реестра (ЕГРЮЛ/ЕГРИП).
Для подготовки такого заявления необходимо зайти в программе «СТЭК — ТРАСТ» меню «ФНС» — «Письма» — «Подготовить заявление на выписку из Единого государственного реестра».
Меню «ФНС» — «Письма» —
«Подготовить заявление на выписку из Единого государственного реестра»
В данном случае открывается формирование письма с шаблоном заявления на выписку из ЕГРЮЛ/ЕГРИП. Программа проверяет корректность заполнения банковских реквизитов налогоплательщика и в случае их отсутствия (ошибок заполнения) появится окно с предложением перейти к их редактированию.
Программа проверяет корректность заполнения банковских реквизитов налогоплательщика и в случае их отсутствия (ошибок заполнения) появится окно с предложением перейти к их редактированию.
Окно «Предупреждение» о возможности проблем в заполнением реквизитов
Для автоматического заполнения шаблона заявления обязательно необходимы заполненные банковские реквизиты налогоплательщика и ФИО, должность, ИНН руководителя (предпринимателя для физических лиц и индивидуальных предпринимателей).
В случае корректности заполнения реквизитов в программе открывается окно подготовки запроса на выписку из Единого государственного реестра со следующими полями:
•Налогоплательщик – выбирается из выпадающего списка по кнопке , который формируется на основе справочника «Абоненты Системы».
•Сертификат ЭП — данные из сертификата, которым будет подписано заявление.
•Налоговый орган по месту учета – выбирается из списка доступных направлений сдачи отчета (см. справочник «Абоненты Системы» — раздел «Направления предоставления отчетности в гос.
•Налоговый орган по месту нахождения — выбирается из списка доступных направлений сдачи отчета (см. справочник «Абоненты Системы» — раздел «Направления предоставления отчетности в гос. органы») по кнопке .
•Файл со скан-копией платежного поручения – необходимо приложить скан-копию платежного поручения (об оплате за предоставление сведений из Единого государственного реестра).
•Тема письма – автоматически прописывается текст «Запрос на выписку из Единого государственного реестра» для идентификации письма в «Реестре писем» и корректировке не подлежит.
•Текст письма – заполняется автоматически шаблон заявления с указанием банковских реквизитов налогоплательщика (фирменный бланк).
Окно «Запрос на выписку из Единого государственного реестра»
После заполнения всех данных нажимается кнопка «Подготовить к отправке». В случае, если не указан файл со скан-копией платежного поручения программа выдаст предупреждение.
Окно «Предупреждение» об отсутствии скан-копии платежного поручения
В зависимости от настроек параметров системы (автоподпись документов) в программе «СТЭК — ТРАСТ» появится одно из двух сообщений о готовности письма к отправке.
Подтверждение формирования письма
После подготовки запроса на выписку из ЕГРЮЛ, он будет находиться в «Реестре писем ФНС».
Если письмо не было подписано автоматически, то «Реестре писем» по кнопке «Подписать» необходимо поставить ЭП на письмо с заявлением, которое требуется отправить. При выходе на связь по кнопке «Отправить / получить» подписанное ЭП письмо будет отправлено в налоговый орган. Процедура отправки писем описана в разделе «Письма» данного Руководства.
Письма
Банки смогут запрашивать в ФНС выписки о доходах с согласия клиентов » Регуляторы
Данные изменения содержатся в проекте ЦБ о внесении изменений в Указание Банка России от 31 августа 2018 года № 4892-У «О видах активов, характеристиках видов активов, к которым устанавливаются надбавки к коэффициентам риска, и методике применения к указанным видам активов надбавок в целях расчета кредитными организациями нормативов достаточности капитала».
В пояснительной записке к документу говорится, что в целях повышения оперативности получения и актуальности документов, получаемых банками из государственных информационных сервисов, предлагается предоставить им право использовать при определении величины среднемесячного дохода заемщика доходы, подтвержденные сведениями о выплатах, произведенных плательщиками страховых взносов в пользу физических лиц, полученными из информационной системы ФНС России.
В Банке России порталу PLUSworld.ru рассказали, что данные изменения в Указание № 4892-У внесены по поручению Председателя Правительства РФ и направлены на обеспечение возможности оперативного получения кредитными организациями сведений ФНС России о доходах заемщика посредством сервиса СМЭВ. В рамках реализации этого Поручения ФНС России при участии Банка России разрабатывает формат и порядок представления указанных сведений по запросам кредитных организаций с получением обязательного согласия заемщика.
В настоящее время уже реализована возможность получения банками информации о выплатах, произведенных плательщиками страховых взносов в пользу физических лиц, из Пенсионного фонда РФ (ПФР) посредством Единого портала государственных и муниципальных услуг с использованием сервиса СМЭВ «Интеграционный модуль» с получением обязательного согласия заемщика на получение банком выписки ПФР. При этом сведения о доходах физических лиц в выписках ПФР в настоящее время обновляются с периодичностью до шести месяцев.
Как рассказали в ВТБ, банкиры давно обсуждают необходимость дополнительных источников информации: «Это нужно для повышения объективности расчета ПДН, что приведет к росту доступности кредитования и улучшению клиентского пути».
По мнению экспертов, успешная реализация идеи зависит от того, будет ли ФНС готова делиться данными и на каких условиях. Правозащитники видят в предложенном ЦБ подходе скорее плюсы для заемщиков: это даст банкам возможность видеть реальную картину доходов граждан.
По материалам Банка России, Коммерсантъ
На сайте ФНС России можно бесплатно получить выписку из ЕГРЮЛ / ЕГРИП о конкретном юридическом лице
Версия для печати 08.2015
08.2015
ФЕДЕРАЛЬНАЯ НАЛОГОВАЯ СЛУЖБА
ИНФОРМАЦИЯ
от 25 июня 2015 года
Важнейшей задачей налоговых органов является развитие интерактивных сервисов, позволяющих налогоплательщикам получать государственные услуги в любое удобное время, в любом месте, используя сеть Интернет.
В этой связи, ФНС России постоянно расширяет спектр оказываемых в электронном виде государственных услуг, а также улучшает качество их предоставления.
Так, например, на сайте ФНС России функционирует модернизированный интерактивный сервис «Предоставление сведений из ЕГРЮЛ/ЕГРИП о конкретном юридическом лице/индивидуальном предпринимателе в форме электронного документа», позволяющий всем заинтересованным лицам бесплатно получать сведения из Единого государственного реестра юридических лиц/Единого государственного реестра индивидуальных предпринимателей в виде выписки/справки об отсутствии запрашиваемой информации в форме электронного документа в формате pdf, подписанного электронной подписью.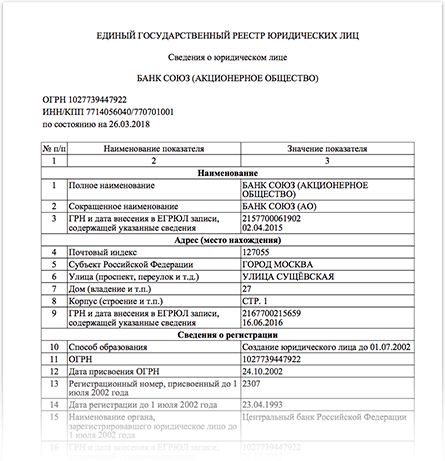
Для получения выписки/справки об отсутствии запрашиваемой информации в форме электронного документа не требуется сертификат ключа электронной подписи (СКП) заявителя.
Для регистрации в сервисе достаточно указать адрес электронной почты, на который поступит ссылка для скачивания выписки.
Сформированную выписку/справку можно скачать в течение 5 дней.
Справка для ФНС об оплате медицинских услуг ИНВИТРО
Информация о порядке предоставления справок об оплате медицинских услуг для представления в налоговые органы Российской Федерации
В соответствии с подпунктом 3 пункта 1 статьи 219 Налогового кодекса РФ налогоплательщик имеет право на получение социального налогового вычета в размере суммы, уплаченной им в налоговом периоде за медицинские услуги, оказанные медицинскими организациями ему, его супругу (супруге), родителям, детям (в том числе усыновленным) в возрасте до 18 лет, подопечным в возрасте до 18 лет (в соответствии с перечнем медицинских услуг, утвержденным Правительством РФ), с учетом ограничения по сумме, установленного пунктом 2 статьи 219 Налогового кодекса РФ.

Право на применение социального налогового вычета, предусмотренного подпунктом 3 пункта 1 статьи 219 Налогового кодекса РФ, имеют все плательщики НДФЛ- получатели доходов, облагаемых по ставке 13%.
Справка об оплате медицинских услуг для представления в налоговые органы РФ выдаётся по требованию налогоплательщика, производившего оплату медицинских услуг, и оформляется бесплатно. Справка оформляется в соответствии с Приказом Минздрава РФ N 289, МНС РФ N БГ-3-04/256 от 25.07.2001.
Для получения справки необходимо предоставить документы, подтверждающие произведенные расходы (кассовые чеки, банковские выписки, квитанции, бланки строгой отчетности), и сообщить ФИО налогоплательщика, ИНН налогоплательщика (при его наличии). Срок подготовки справки – до 25 рабочих дней.
Если медицинские услуги оплачены налогоплательщиком для супруга (супруги), родителей или детей, необходимо дополнительно предоставить сведения о ФИО лица, которому были оказаны медицинские услуги, и степени родства с налогоплательщиком. В справке сведения указываются со слов налогоплательщика. В налоговый орган подается копия документа, подтверждающего степень родства (например, свидетельство о рождении, свидетельство о браке), налоговый орган вправе запросить оригинал документа.
В справке сведения указываются со слов налогоплательщика. В налоговый орган подается копия документа, подтверждающего степень родства (например, свидетельство о рождении, свидетельство о браке), налоговый орган вправе запросить оригинал документа.
Порядок оформления справки уточняйте у администраторов Медицинских офисов и по телефонам справочно-информационной службы 8 (495) 363-0-363 (для звонков из Москвы), 8 (800) 200-363-0 (для звонков из регионов, звонок по России бесплатный).
Внимание! Уважаемые пациенты! Справка установленного образца выдается только при наличии документов, подтверждающих произведённые расходы. Обращаем Ваше внимание, утерянные, выцветшие кассовые чеки не восстанавливаются.
Инспекция ФНС России № 50
ИФНС 50
Начальник Инспекция ФНС России № 50:
Феликсова Наталия Михайловна
Фактический адрес:
125373, г. Москва, Походный проезд, вл.3, корп.А
Юридический адрес:
125373, г. Москва, Походный проезд, вл.3, корп.А
Москва, Походный проезд, вл.3, корп.А
Адрес для почтовых отправлений:
125373, г. Москва, Походный проезд, вл.3, корп.А
Схема проезда до налоговой инспекции:
Cпособ проезда:
Станция метро «Сходненская», 1-й вагон из центра, маршрутное такси № 368 или последний вагон из центра, авт. № 678, № 199; станция метро «Тушинская», последний вагон из центра, авт. № 2, № 266 до остановки «Платформа Трикотажная», автобус №88, №777 до остановки «17 Таксомоторный парк»
Телефоны:
Контакт-центр: 8-800-222-22-22
Приемная начальника Инспекции: 8 (495) 400-00-50
Для справок: 8 (495) 400-37-15
Факс:8 (495) 400-37-11
Отдел | |
|---|---|
Отдел урегулирования задолженности и обеспечении процедур банкротства | |
Отдел работы с налогоплательщиками | |
Отдел заказа выписки |
Реквизиты
ИНН 7702265064
КПП 773301001
Код налогового органа в ФНС 50: 7750
Получатель: Управление Федерального казначейства по г. Москве (ИФНС России №50 по г. Москве)
Москве (ИФНС России №50 по г. Москве)
Наименование банка: ГУ Банка России по ЦФО
Номер счета: 40101810045250010041
БИК банка: 044525000
Номер корреспондентского счета или субсчет банка в котором открыт счет УФК России по г. Москве – нет
| Время pa6oты ИФНС: | Пepepыв: | |
пн: | 9.00-18.00 | 13.00-13.45 |
вт: | 9.00-18.00 | 13.00-13.45 |
cp: | 9.00-18.00 | 13.00-13.45 |
чт: | 9. 00-18.00 00-18.00 | 13.00-13.45 |
пт: | 9.00-16.45 | 13.00-13.45 |
Прием налогоплательщиков в операционном зале осуществляется без перерыва на обед понедельник, среда с 9.00 до 18.00; | ||
Выдача готовых выписок из ЕГРЮЛ/ЕГРИП При доставке выписки из ЕГРЮЛ/ЕГРИП, время согласовывается отдельно. | ||
<< Назад к списку ИФНС Москвы
Если Вы заметили на сайте опечатку или неточность, выделите её |

Выписка из ЕГРН налоговая. Проверка по ЕГРН на сайте налоговой
Получение налогового вычета за имущество требует предоставления документов на имущество. Что нужно знать о налоговой выписке из ЕГРН?
Когда требуется выписка из ЕГРН? Налоговая служба требует эту бумагу при совершении сделок с имуществом. Но как ее получить и что это за документ?
Выписка из ЕГРН для налогового вычета – для чего нужна
Для начала разберемся, для каких целей оформляется изучаемая документация.
Выписка из ЕГРН для налогового вычета необходима преимущественно при приобретении недвижимости. Возврат средств при продаже собственности тоже имеет место, но не всегда.
Выписка из ЕГРН налоговая — как выглядит
В любом случае, выписка из ЕГРН (налоговая) запрашивается,если:
— гражданин продал имущество или сдал его;
— произошла покупка недвижимости.
Представляют соответствующую справку об объекте недвижимости при подаче декларации о доходах или во время запроса вычета имущественного типа. Без причин ФНС не может требовать упомянутую бумагу.
Без причин ФНС не может требовать упомянутую бумагу.
Выписка из ЕГРН налоговая – содержание документа
Какие сведения можно увидеть в налоговой выписке из ЕГРН? Это самые обычные данные из реестра недвижимости. И они ничем не отличаются от общепринятых сведений из Госреестра.
То есть, в выписке из ЕГРН для налогового вычета собственник увидит:
— основание выдачи бумаги;
— характеристики недвижимости;
— Ф. И. О. собственников;
— наличие ограничений и обременений;
— сведения о регистрирующем органе.
Если запросить расширенный тип справки, можно дополнительно просмотреть историю перехода собственности от человека к человеку. Но ФНС подобной информацией почти не интересуется. Налоговой достаточно обычной выписки с характеристиками объекта. На ней мы остановимся более подробно.
Выписка из ЕГРН для налогового вычета – где можно запросить
Немаловажно понимать, где оформляется изучаемые бумаги. От этого будут зависеть дальнейшие действия по получению справки из реестра недвижимости РФ.
На сегодняшний день выписка из ЕГРН может быть получена в МФЦ.
Справка ЕГРН налоговая – как получить самостоятельно
Для начала рассмотрим алгоритм действий при запросе сведений из Госреестра лично. Это самый простой вариант развития событий.
Если собственность только приобретена, то с выданным при заключении сделки пакетом документов гражданин должен обратиться в один из регистрирующих органов. Спустя некоторое время, человеку выдадут справку установленного образца. В противном случае нужно подать запрос с документами, оплатить услугу и ждать.
После проделанных действий налоговая инспекция заберет у собственника выписку и учтет ее во время рассмотрения запроса по возврату средств за покупку недвижимости. Более подробную информацию по оформлению вычетов можно получить на сайте nalog.ru.
Справка ЕГРН налоговая – оформляем онлайн через Госреестр
В 2018 году в бумажном или электронном виде можно получить выписку из ЕГРН для налогового вычета в режиме онлайн. К примеру, через официальный сайт.
К примеру, через официальный сайт.
Налоговая выписка ЕГРН для юрлиц
Для того, чтобы подать запрос установленной формы, придется:
1. Зайти на сайт Росреестра.
2. Оформить запрос в электронном виде (заглянуть в «Сервисы»-«Сведения из ГКН»).
3. Оплатить услугу.
Сроки оказания услуги по заказу выписки ЕГРН
Как долго ожидать готовности изученной справки? Ответ зависит от способа подачи запроса.
Выписка из ЕГРН для налоговой с планом квартиры
Пероральная доставка олигомерных процианидинов в Apple Poly® усиливает ответы IFN типа I in vivo
Abstract
Передача сигналов IFN типа I является центральным путем, который обеспечивает критическую врожденную защиту от вирусных и бактериальных инфекций и может иметь регуляторные последствия при воспалительных процессах. Ранее мы определили, что OPC, содержащиеся в диетической добавке APP, усиливали реакцию на IFN типа I in vitro. Здесь мы подтверждаем, что OPC из двух разных источников значительно увеличили pSTAT1, в то время как мономерная форма процианидина этого не сделала.Мы предположили, что подобные ответы могут быть вызваны in vivo после приема APP. Проглатывание APP перед инъекцией polyI: C усиливало in vivo ответы на IFN типа I у мышей. Когда люди принимали АРР, были обнаружены усиленные ответы на IFN типа I и усиленный pSTAT1 ex vivo, тогда как прием RES, мономерного полифенола, индуцировал минимальные такие изменения. Полифенолы наиболее известны тем, что вызывают противовоспалительные и антиоксидантные реакции; тем не менее, наши результаты свидетельствуют об уникальном, неантиоксидантном аспекте OPC, который широко применим ко многим заболеваниям.Способность пероральных OPCs усиливать передачу сигналов IFN типа I in vivo может усиливать врожденную защиту и может частично способствовать отмеченному противовоспалительному результату приема OPC из многих источников.
Ключевые слова: врожденный противовирусный, аутоиммунный, дополнительная терапия
Введение
Передача сигналов IFN типа I является центральным клеточным процессом с широким диапазоном последующих эффектов. IFN типа I индуцируются сразу после обнаружения вирусных молекулярных паттернов и сигнала для защиты инфицированных и неинфицированных клеток, таким образом, они являются основным компонентом врожденного противовирусного иммунитета [1].Таким образом, интерфероны типа I используются для лечения хронических вирусных инфекций [2], и было показано, что они являются эффективными средствами лечения широкого спектра вирусных угроз [3, –6]. IFN типа I также может вызывать противовоспалительную передачу сигналов и долгое время использовался в качестве терапии для пациентов с рассеянным склерозом [7], но только недавно были обнаружены специфические механизмы защиты [8, 9]. IFN типа I также играют решающую роль в раннем противоопухолевом иммунитете за счет стимуляции DC [10] и используются в качестве терапии рака [11]. Кроме того, передача сигналов IFN типа I способствует оптимальному ответу IFN (IFN-γ) типа II [12] и способствует гематопоэзу [13].Передача сигналов IFN типа I, несомненно, важна при множественных заболеваниях и является клинической мишенью. Перорально вводимая новая иммунотерапия, которая усиливает этот сигнальный путь, может иметь широкое терапевтическое применение в качестве комбинированной терапии или потенциально может предотвратить инъекции интерферона.
Полифенолы — это химические соединения, которые в различных концентрациях и типах обнаруживаются во многих пищевых источниках, таких как гранаты, виноград и чай. Широко показано, что полифенолы обладают антиоксидантной и противовоспалительной способностью [14, 15].APP — это экстракт незрелой яблочной кожуры, содержащий высокие концентрации OPCs [16]. Эта добавка обладает заметными противовоспалительными эффектами на модели колита у мышей, которые зависят от αβ Т-клеток [14]. OPC состоят из двух — 10 повторов гидроксилированных строительных блоков флаван-3-ола, известных как катехин или эпикатехин ( ). Уже известно, что ОРС из кожуры яблони и других источников, таких как виноградные косточки и кора морской сосны (пикногенол), безопасны для употребления в пищу [17, 18]. Недавно мы определили способность OPCs из APP усиливать врожденный иммунитет, что явно отличается от антиоксидантной активности [16, 18, –21].Используя in vitro инфицированные вирусом денге человеческие клетки, мы обнаружили, что лечение ОРС, полученными из АРР, снижает вирусные титры и проявляет уникальный противовирусный механизм: усиление ответов на IFN типа I [21]. Здесь эти результаты были подтверждены демонстрацией того, что два надежных источника OPC усиливали фосфорилирование STAT1, которое происходит после взаимодействия IFNR типа I, тогда как мономерный процианидин этого не делал. Основная цель этого исследования заключалась в том, чтобы продемонстрировать, что усиление передачи сигналов IFN типа I можно также измерить in vivo после приема APP.Прием APP усиливал передачу сигналов IFN типа I у мышей, которым вводили дцРНК-миметик polyI: C, и оптимальные ответы были обнаружены через 1 неделю после приема APP. Подчеркивая медицинскую значимость этих данных, прием АРР людьми индуцировал усиленные ответы на IFN типа I в клетках крови, культивируемых ex vivo. Эти данные показывают, что минимальные пероральные дозы АРР усиливают ответы IFN типа I in vivo. Способность простой и безопасной пероральной добавки усиливать реакцию на IFN типа I может иметь широкий потенциал для усиления врожденной защиты от вирусной инфекции или для использования в качестве комбинированной терапии для пациентов, получающих rIFN типа I по поводу хронической вирусной инфекции, аутоиммунных состояний или рака. .
ОРС, а не мономерные процианидины увеличивают pSTAT1.(A) Химическая структура OPC и мономерных процианидинов. (B) Инкубация с надежными источниками OPC (экстракты APP и AS), но не с катехином в культуре, привела к прямому увеличению pSTAT1 (на клетках CD11b + ) с экзогенным IFN типа I и без него. Вставка представляет собой репрезентативный график сравнения pSTAT1 в человеческих PBMC, обработанных OPC, полученными из APP (темная линия) и только катехина и среды (светлые линии).На правой стороне графика клетки сначала инкубировали в течение 24 или 3 часов с APP, затем промывали и инкубировали еще 48 часов, всего 72 часа. Более длительная инкубация усиливала ответы на экзогенный IFN типа I, но устраняла прямые ответы APP на pSTAT1. MFI — средняя интенсивность флуоресценции; Конц., Концентрации. Звездочки обозначают существенное отличие от образцов, содержащих только среду, с аналогичной обработкой. Тест Стьюдента t , непарный (* P <0,05; ** P <0.01; *** P <0,001).
МАТЕРИАЛЫ И МЕТОДЫ
Клетки человека in vitro
Для оценки эффектов OPCs и катехина на pSTAT1 были выделены полные PBMC с использованием Histopaque (Sigma, Сент-Луис, Миссури, США), как описано ранее [21], и культивировали в течение 24 часов с OPC, полученными из APP [21], водным экстрактом APP, недавно идентифицированным источником OPC (метанольный экстракт AS), катехином или только средой. Затем клетки стимулировали в течение 15 минут rIFN-α (100 Ед / мл; человеческий интерферон-альфа A / D; PBL Interferon Source, Piscataway, NJ, USA), фиксировали, пермеабилизировали, окрашивали антителом Phosflow pSTAT1 PE (BD Biosciences , Сан-Хосе, Калифорния, США), используя протоколы производителя, и проанализировали с помощью цитометра FACSCaliber (BD Biosciences) и программного обеспечения FlowJo (Ashland, OR, США).В дополнительных экспериментах клетки обрабатывали в течение 3 или 24 часов APP, затем промывали и инкубировали еще 48 часов в культуре с последующим добавлением rIFN-α и аналогичными анализами.
Индукция ответа IFN типа I путем инъекции полиI: C
Все исследования на животных проводились в соответствии с Комитетом по уходу и использованию животных Университета штата Монтана. Шестинедельным мышам BALB / c давали воду или экстракт АРР в дозах (5 мг / 100 мкл; на основе приблизительной эквивалентности дозе 1 г для человека) [22] с различными интервалами, а затем вводили 1 мкг полиI: C. доставлено i.п. инъекция. Через 4 часа мышей умерщвляли, печень (одну долю) и целые селезенки гомогенизировали, эритроциты лизировали водой и клетки лизировали для выделения РНК с использованием набора RNeasy Mini Kit (Qiagen, Валенсия, Калифорния, США).
Исследования на людях
Исследования на людях проводились в соответствии с Комитетом по надзору за учреждениями Университета штата Монтана, и поэтому каждый субъект подписал документацию об информированном согласии. В первом исследовании кровь была взята у добровольцев, которые ничего не принимали, или у тех же субъектов ( n = 11), через 2 недели, от 90 минут до 2 часов после приема 3 г APP (шесть капсул; Apple Poly®, Morrill , NE, США).Этот чередующийся образец повторялся четыре раза (раунды 1–4) и продолжался в отсутствие приема APP в течение трех дополнительных циклов (раунды 5-7). Во втором исследовании прием APP сравнивался с приемом RES в рандомизированном и двойном слепом режиме. Начиная с 10 субъектов / группу, группы принимали 3 г АПП или 3 г РЭС (Биотивиа Трансмакс), а кровь собирали через 2 часа (Раунд 1). Второй раунд был повторен через 3 недели. Раунд 3, в котором субъекты не принимали добавки, был проведен через 4 недели, и были сравнены те же параметры.В обоих исследованиях группы субъектов чередовались и смешивались таким образом, чтобы кровь максимум шести субъектов анализировалась в день, чтобы уменьшить возможные эффекты ежедневных вариаций, искажающих результаты. Других ограничений в питании и деятельности волонтеров не было. Полные PBMC выделяли, как описано ранее [21], и стимулировали ex vivo средой или 100 ед / мл rIFN-α. Клетки в отдельных лунках стимулировали в течение 4 часов, а затем лизировали для выделения РНК и КПЦР или культивировали в течение 24 часов для анализа методом проточной цитометрии.Индукцию складки рассчитывали делением значения, обработанного IFN, на значение только среды для клеток от того же донора.
Проточная цитометрия с клетками человека
PBMC человека окрашивали антителами, специфичными для CD11b (для идентификации мононуклеарных клеток; клон ICRFF44; BD PharMingen, Сан-Диего, Калифорния, США) и MHC I (клон W6 / 32; BioLegend, Сан-Диего , CA, США) и CD86 (Clone IT2.2; BioLegend), экспрессия которых увеличивается в ответ на IFN типа I [23, 24]. Клетки окрашивали с использованием стандартных протоколов проточной цитометрии и анализировали с помощью цитометра FACSCaliber.Данные анализировали с помощью программного обеспечения FlowJo, MS Excel (Microsoft, Редмонд, Вашингтон, США) и Prism (GraphPad Software, Ла-Холья, Калифорния, США).
Экстракция РНК и RT-qPCR
РНК из тканей мыши и клеток человека экстрагировали, подвергали обратной транскрипции, а также оценивали и анализировали уровни транскриптов, как описано ранее [21].
Статистический анализ
Статистическая значимость определялась с помощью параметрических и непараметрических тестов. Представленные данные были проанализированы с использованием непарного одностороннего теста t , если не указано иное.
РЕЗУЛЬТАТЫ И ОБСУЖДЕНИЕ
Для подтверждения ранее полученных данных in vitro и исследования того, является ли действие АРР на путь IFN типа I специфичным для соединений, содержащих OPC, был проведен анализ pSTAT1 in vitro. Человеческие PBMC инкубировали в течение 24 часов с водным экстрактом APP, водным экстрактом другого недавно идентифицированного надежного источника OPC (AS), OPC, полученных из APP, как описано ранее [21], катехина или только среды. B демонстрирует, что более низкие концентрации APP и AS увеличивают pSTAT1 на клетках CD11b + в ответ на rIFN-α.После 24 ч инкубации AS при 1 мкг / мл и все источники OPC при 10 мкг / мл, включая OPC, выделенные из APP (B, вставка), также индуцировали pSTAT1 в отсутствие rIFN-α. Инкубация в течение 1, 3 или 6 часов с 10 мкг / мл АРР не вызывала таких изменений (данные не показаны). Катехин, мономерный процианидин, вызывал ответы, подобные контрольным, содержащим только среду. Эти данные предполагают, что OPCs и источники OPCs, но не мономерные процианидины, увеличивают pSTAT1 in vitro и что мономерные полифенолы могут использоваться в качестве отрицательного контроля.Дополнительные эксперименты касались необходимого интервала для предварительной обработки APP. Чтобы исследовать более длительный ответ, клетки обрабатывали АРР в течение 3 или 24 часов, затем промывали и инкубировали еще 48 часов перед обработкой rIFN-α. В этом случае прямой pSTAT1 в отсутствие rIFN-α не пострадал, но в ответ на rIFN-α pSTAT1 был значительно увеличен обработкой APP по сравнению с контролями только со средой. Эти данные предполагают, что лечение APP заставляет клетки лучше реагировать на IFN типа I, и для полного развития максимального примирования ответа требуется несколько дней.Противовоспалительные эффекты — это наиболее часто описываемый эффект от приема полифенолов [14, 15], который, как правило, может быть связан с их антиоксидантной способностью. OPCs, но не мономерные формы, снижают титры вируса денге in vitro [21] и повышают pSTAT1, даже несмотря на то, что эти полифенолы обладают аналогичной антиоксидантной способностью. В совокупности наши результаты показывают, что OPC обладают новым и уникальным действием на ответы на передачу сигналов IFN типа I, которое отличается от хорошо изученных антиоксидантных эффектов.
Эффекты приема экстрактов АРР на ответы на IFN типа I in vivo оценивали путем измерения экспрессии транскриптов ISG после инъекции мышей polyI: C.Для определения оптимальной пероральной дозы АРР мышей разделили на четыре группы лечения и получили через желудочный зонд одну дозу 5 мг АРР за 36 ч до инъекции; четыре дозы по 5 мг АРР; или воду только за 36, 24, 12 и 0 ч до инъекции, или те же четыре дозы каждые 12 ч за 1 неделю до инъекции. Мышам инъецировали polyI: C, и ткани собирали через 4 часа и обрабатывали для анализа RT-qPCR. Четыре часа были эмпирически определены как оптимальная точка для экспрессии транскрипта ISG после инъекции polyI: C.Транскрипты ISG не были обнаружены ни после приема одного АРР, ни через 1 час после инъекции полиI: C (данные не показаны). Транскрипты ISG SOCS1 , OAS1 , IFIT1 и IFIT2 индуцировались более устойчиво в селезенке, тогда как экспрессия CXCL10 была выше в печени ( A ). В соответствии с данными in vitro, предполагающими повышенную активность после нескольких дней (48 ч) инкубации, доставка APP за 1 неделю до инъекции polyI: C была наиболее эффективным интервалом дозирования для усиления ответа IFN типа I, поскольку эта доза значительно увеличивала экспрессию SOCS1. , IFIT1 , IFIT2 и CXCL10 по сравнению с контролями с подачей воды.Только четыре дозы в течение 36 часов после инъекции значительно изменили экспрессию OAS1 и в печени, а кормление APP не изменило экспрессию OAS1 и в селезенке. Четыре дозы APP в течение 36 часов после инъекции также значительно повлияли на экспрессию IFIT1 и IFIT2 в печени, так же как одна доза, 36 часов назад, повлияла на экспрессию IFIT2 . Доставка APP за 1 неделю до инъекции вызвала кратное увеличение на 4,7 и 3,9 раза экспрессии CXCL10 в селезенке и печени, соответственно, по сравнению с мышами, получавшими воду.Это увеличение было намного больше, чем индукция других ISG, которая составляла от 1,8 до 2,8 раза. Эти данные показывают, что пероральный APP влиял на экспрессию ISG in vivo, и интервал дозирования, который больше всего влиял на экспрессию гена, составлял 1 неделю до инъекции polyI: C. Относительно долгосрочный эффект минимального приема внутрь OPC согласуется с данными in vitro и может указывать на то, что OPCs примировали клетки для усиления ответа на IFN типа I in vivo, а для оптимального обнаружения примированного состояния в большой фракции клеток требуются дни, чтобы полностью развивать.
Пероральный APP усиливал ответы на IFN типа I in vivo.Мышей ( n = 5) кормили 5 мг дозами APP или водой с различными интервалами, как указано на оси x , а затем вводили 1 мкг polyI: C. (A) Четыре дозы APP за 1 неделю до инъекции polyI: C были наиболее эффективной дозой для усиления экспрессии ISG в селезенке и печени, хотя другие интервалы дозирования были эффективны для повышения экспрессии некоторых ISG. (B) После четырех доз APP за 1 неделю до инъекции полиI: C транскрипты IFN типа I были более детектируемыми в печени, чем в селезенке; однако существенные различия между мышами, получавшими APP, и мышами, получавшими воду, были обнаружены только в селезенке для IFN-α (* P <0.05; ** P <0,01; *** P <0,001 по непарному t -тест; звездочки обозначают статистическую значимость разницы между указанной группой и группой, обработанной водой).
Инъекция PolyI: C индуцирует экспрессию IFN I типа; однако потенциальной причиной повышенной экспрессии ISG в группе, получавшей APP, может быть дальнейшее усиление экспрессии самих IFN типа I, поэтому эта возможность была исследована. Транскрипты, кодирующие IFN типа I, измеряли в селезенке и печени мышей, получавших воду или APP с наиболее эффективным интервалом дозирования, за 1 неделю до инъекции polyI: C (B).Транскрипты IFN типа I обнаруживались после инъекции полиI: C, и уровни были одинаковыми в двух группах. По сравнению с печенью транскрипты IFN типа I обнаруживались только на очень низких уровнях в селезенке. Единственная значимая разница между группами, получавшими воду и АРР, заключалась в 1,65-кратном увеличении экспрессии IFN-α1 в селезенках группы, получавшей АРР, по сравнению с мышами, получавшими воду. Не было отмечено значительных различий между группами в печени для транскриптов IFN-α1 или IFN-β (B).Подобным образом и как ожидалось, не было различий ни в одной из тканей в экспрессии транскриптов IFN-γ (данные не показаны). Несколько более высокая экспрессия IFN-α1 в селезенке мышей, получавших APP, предлагает возможное объяснение увеличения экспрессии ISG, наблюдаемого в этой ткани, но это явно не имело место в печени, где ISG увеличивались независимо от IFN типа I. выражение. Эти данные подтверждают наши результаты in vitro и позволяют предположить, что прием OPCs существенно не изменяет экспрессию IFN I типа, но усиливает передачу сигналов IFN I типа in vivo.
Чтобы максимизировать клинический и трансляционный потенциал наших результатов, были проведены небольшие эксперименты на человеке с субъектом, чтобы продемонстрировать, что прием АРР усиливает ответы на IFN типа I у человеческих PBMC, культивируемых ex vivo. В первом исследовании здоровые субъекты ничего не потребляли (NEG), а через 2 недели потребляли 3 г APP, и эта схема была повторена. Ответы сравнивали с PBMC от того же человека; таким образом, каждый человек служил своим собственным отрицательным контролем. Через 24 часа в культуре ex vivo и без какого-либо другого лечения наблюдалось значительное увеличение средней флуоресценции CD86 после первой и второй доз АРР по сравнению с тем, когда те же субъекты изначально ничего не принимали.В начале раунда 3 экспрессия CD86 оставалась значительно повышенной по сравнению с уровнями в начале раунда 1 и не увеличивалась далее в последующих раундах при приеме APP ( A ). Таким образом, несмотря на прекращение приема АРР, экспрессия CD86 оставалась повышенной в течение нескольких недель, что свидетельствует об очень длительном влиянии на экспрессию CD86, вызванном множественными дозами АРР. Аналогичная картина была очевидна для индукции MHC I в ответ на обработку rIFN-α ex vivo.Кратность индукции MHC I (на клетках CD11b + ; B) была значительно выше после приема первого и второго APP. Индукция MHC I в начале 3-го и 4-го раундов была значительно выше, чем в начале 1-го раунда, и больше не увеличивалась в этих более поздних раундах при приеме добавок. При использовании клеток от тех же субъектов параллельный анализ индуцированной экспрессии генов в ответ на rIFN-α показал аналогичную тенденцию, наблюдаемую с MHC I. Мы измерили транскрипты ISG: IFIT1 , MX1 , SOCS1 , и RIGI после обработки IFN типа I в течение 4 часов.ISG не экспрессировались в отсутствие лечения IFN типа I. RIGI последовательно индуцировался IFN типа I, и индукция кратности (по сравнению с необработанными клетками того же донора) увеличивалась после приема APP (C). Как и в случае с MHC I и CD86, последующие раунды больше не индуцировали экспрессию RIGI , и его кратная индукция в начале раундов 3 и 4 без приема АРР (NEG) была так же увеличена по сравнению с началом раунда 1. Индукция IFIT1 , SOCS и MX1 не был таким согласованным, как RIGI (данные не показаны).Эти данные показывают, что прием минимальных доз богатого источника OPCs может усиливать экспрессию белков и транскриптов ex vivo, индуцированную низкими уровнями IFN типа I, и что эффект нескольких доз может быть длительным.
Проглатывание полифенолов людьми усиливало ответы ex vivo на IFN типа I.Люди ( n = 11) ничего не принимали или спустя 2 недели однократную дозу 3 г АРР. Это повторялось в течение четырех раундов. Во время 5-7 раундов (R) прием АРР был прекращен, и кровь собиралась у тех же субъектов с 2-недельными интервалами.(A) Экспрессия CD86 на PBMC увеличивалась после приема АРР и оставалась повышенной после нескольких доз. (B) Кратковременная индукция MHC I (на клетках CD11b + , нормализованных только для среды) при 24-часовой обработке rIFN-α увеличивалась во время чередующихся 2-недельных интервалов приема NEG и APP и оставалась повышенной. (C) Индукция транскриптов RIGI при 4-часовой инкубации с IFN-α была увеличена в 1-м раунде и оставалась повышенной после 2-го раунда (в паре, * P <0,05; ** P <0.01; *** P <0,001).
Во втором исследовании прием АРР сравнивали непосредственно с приемом такой же дозы РЭС, и после приема двух доз каждого из них с интервалом 3 недели между дозами наблюдались значительные различия между группами. Наблюдалось значительное увеличение кратности индукции MHC I в клетках, полученных от тех, кто принял две дозы APP, по сравнению с теми, кто принял такое же количество RES ( A ). Однако в этом исследовании не было различий в экспрессии CD86 между субъектами, принимавшими APP, и субъектами, принимавшими RES; скорее, средняя флуоресценция CD86 увеличилась в обеих группах.Эти данные предполагают, что экспрессия CD86 может быть увеличена при приеме полифенолов в целом или повышена эффектом плацебо. Подобно результатам для MHC I, кратная индукция экспрессии RIGI была значительно выше после приема APP по сравнению с приемом RES (B). Мы расширили это исследование, включив в него транскрипты, кодирующие IL-10 и IL-27, так как эти противовоспалительные цитокины, вероятно, участвуют в пользе лечения IFN типа I при экспериментальном аутоиммунном энцефаломиелите, модели РС на мышах [8, 9] .Оба этих транскрипта также индуцировались на значительно более высоком уровне у субъектов, принимавших АРР, по сравнению с субъектами, принимавшими RES (B). Противовоспалительные реакции после приема некоторых полифенолсодержащих добавок могут быть частично результатом усиленных ответов IFN типа I за счет повышенной экспрессии противовоспалительных цитокинов IL-10 и IL-27. В частности, недавно было показано, что передача сигналов IL-27R специфически снижает экспрессию воспалительных цитокинов, в первую очередь IFN-γ, и трафик CD4 + Т-клеток [25, 26].Эти клетки являются основным источником воспаления в модели колита у мышей [27]. APP-индуцированная экспрессия IL-27 посредством усиленной передачи сигналов IFN типа I может быть частично ответственной за отмеченное подавление воспаления с помощью APP, которое зависело от αβ Т-клеток в модели колита [14]. Повышение экспрессии IFN типа I может быть нераспознанным новым механизмом, ответственным по крайней мере за некоторые из отмеченных положительных и иммуномодулирующих результатов после приема пищевых добавок, содержащих OPC.Из-за плейотропных эффектов IFN типа I этот ответ может быть замаскирован в других исследованиях устойчивыми антиоксидантными и ингибирующими ответами, общими для полифенолов, или ответы действительно являются уникальным аспектом OPC из ограниченных источников.
Прием АРР индуцировал усиленные ответы на IFN типа I по сравнению с приемом RES.Субъекты приняли две дозы 3 г АРР ( n = 7) или 3 г RES ( n = 9) с интервалом в 3 недели. (A) Прием APP приводил к усиленной индукции экспрессии MHC I в ответ на IFN типа I по сравнению с приемом RES.Экспрессия CD86 в клетках, обработанных средой, увеличивалась в крови в обеих группах лечения. (B) После двух доз каждой добавки, относительно RES, прием АРР человеческими субъектами приводил к усиленной индукции IFN-стимулированных транскриптов RigI , IL-10 и IL-27 в ответ. к rIFN-α ex vivo. После второго приема АРР или РЕС клетки крови от подгруппы людей собирали и оценивали на pSTAT1. (C) Репрезентативный график FACS, демонстрирующий повышенный исходный уровень (среда, слева) и rIFN-α-стимулированный (справа) pSTAT1 у субъектов, которые принимали APP (заштрихованы), по сравнению с субъектами, которые принимали RES (линии).(D) MFI pSTAT1 указывает на повышенное фосфорилирование у субъектов, принимавших АРР ( n = 4), по сравнению с субъектами, принимавшими RES ( n = 6). Эти данные сравнивали с использованием непарного, t -теста, * P <0,05; ** P <0,01.
Мы также оценили различия в pSTAT1 ex vivo в клетках человека после двух доз APP или RES. PBMC от подгруппы субъектов культивировали в течение ночи, а затем стимулировали rIFN-α в течение 15 минут, фиксировали, пермеабилизировали и окрашивали антителом, специфичным к pSTAT1.Подобно результатам in vitro (B), как базальные, так и стимулированные rIFN-α уровни pSTAT1 были немного увеличены у субъектов, принимавших APP, по сравнению с теми, кто принимал RES (C). Когда этот ответ сравнивали у нескольких доноров, было статистически значимое увеличение IFN-α-индуцированного pSTAT1 у субъектов, которые принимали APP, по сравнению с субъектами, которые принимали RES (D). Для каждого из параметров, показанных в, не было значительных различий между группами APP и RES после одной дозы, а также различия не сохранялись через 4 недели после второй дозы (данные не показаны), что позволяет предположить, что необходимы множественные, случайные, небольшие дозы OPC. для сохранения эффектов, а двух доз недостаточно для длительного изменения.Эти данные показывают, что прием АРР индуцировал усиленные ответы IFN типа I у мышей и людей. Этот ответ может зависеть от OPC, обнаруженных в ограниченном наборе пищевых источников.
Пероральная терапия широкого спектра действия, которая усиливает ответы на IFN I типа, ранее не описывалась. IFN типа I индуцируются сразу после определения вирусных молекулярных паттернов и являются критическим компонентом врожденного противовирусного иммунитета. Наша терапевтическая защита от вирусов в настоящее время ограничивается несколькими противовирусными препаратами и лечением rIFN типа I [4].Наши данные свидетельствуют о том, что проглоченный APP оказывает длительное влияние на паттерны экспрессии, расположенные ниже по течению от IFN типа I. Эти результаты подтверждают новый биологический эффект источников OPC in vivo, который может быть жизнеспособным противовирусным профилактическим подходом. RIFN типа I используется в противовирусной терапии вируса гепатита С человека [2, 28] и эффективен на моделях инфекции гриппа [3, 5]. Таким образом, пероральные источники OPC могут также использоваться в качестве комбинированной терапии для усиления эффектов терапии IFN типа I при хронической вирусной инфекции [2, 28], особенно с учетом обнаруженных ответов в печени после приема APP у мышей.Любой человек с повышенным риском вирусной инфекции и осложнений, включая пожилых людей, родителей маленьких детей, учителей и медицинских работников, может получить большую пользу от усиленной врожденной защиты от вирусной инфекции.
IFN типа I также давно используется в качестве терапии для пациентов с РС [7, 29], но только недавно были обнаружены специфические механизмы защиты, включающие усиленную экспрессию противовоспалительных цитокинов IL-10 и IL-27 [8, 9]. Ответы на терапию интерфероном I типа сильно различаются между пациентами; таким образом, пероральное лечение, которое в целом увеличивает ответ на IFN типа I, может повысить эффективность этой терапии.Осложнением лечения IFN типа I является необходимость введения больших непрерывных доз, в результате чего IFN-специфические нейтрализующие антитела делают лечение менее эффективным [29]. Использование пероральных OPC в качестве комбинированной терапии с введенными или принимаемыми внутрь IFNs типа I может потенциально снизить эффективную дозу и, таким образом, уменьшить побочные эффекты. Более того, усиление ответов на эндогенно экспрессируемые IFN типа I может устранить необходимость в какой-либо дополнительной терапии. Для большинства воспалительных состояний, включая рассеянный склероз, доступна только паллиативная терапия; таким образом, изобретательные терапевтические подходы явно оправданы.
Есть несколько аспектов заболевания, на которые не влияет передача сигналов IFN типа I. IFN типа I имеют решающее значение для иммунного ответа на опухоли [10], могут влиять на кроветворение [30] и недавно были признаны решающими для защиты от некоторых бактериальных инфекций [31, –33]. Характеристика этого нового эффекта приема внутрь OPCs на передачу сигналов IFN типа I, вероятно, даст представление о механизмах in vivo и убедительное обоснование для его рекомендованного применения в широком диапазоне болезненных состояний.Принимая во внимание нехватку и осложнения существующих методов лечения и далеко идущие последствия пути IFN типа I, способность простой пероральной добавки усиливать ответы на IFN типа I, экспрессируемого эндогенно во время инфекции или доставляемого в виде терапевтического рекомбинантного белка, может в значительной степени воздействовать на поле.
Интерфероны III типа при вирусных инфекциях и противовирусном иммунитете — FullText — Cellular Physiology and Biochemistry 2018, Vol. 51, № 1
Аннотация
Интерфероны (IFN) могут служить первой линией иммунной защиты от вирусной инфекции.Идентификация IFN-λs 1, 2, 3 и 4 (называемых IFN типа III) показала, что противовирусный иммунный ответ на вирусы содержит больше компонентов, чем IFN типа I, которые известны более 50 лет. IFN-λ представляют собой IFN-λ1 (IL-29), IFN-λ2 (IL-28a), IFN-λ3 (IL-28b) и IFN-λ4, который напоминает IFN-λ3. IFN-λ обладают иммунными ответами и биологической активностью типа I-IFN, но наши знания об этих новых участниках противовирусного ответа недостаточно хорошо установлены. В этом обзоре мы пытаемся описать текущую информацию об экспрессии и функции IFN-λ во врожденной противовирусной иммунной защите и роли IFN-λ2 в регуляции и формировании адаптивного иммунного ответа.Мы предполагаем, что IFN-λ являются ключевыми противовирусными цитокинами, непосредственно осуществляющими противовирусный иммунный ответ на эпителиальных поверхностях на ранних стадиях вирусной инфекции, и что эти цитокины также искажают баланс клеток Th2 и Th3 в сторону фенотипа Th2. Кроме того, генетический полиморфизм генов IFN-λ может ухудшать противовирусные иммунные ответы при клиническом лечении.
© 2018 Автор (ы). Опубликовано S. Karger AG, Базель
Введение
Первая линия защиты для противодействия патогенным инфекциям обычно зависит от врожденного иммунного ответа.В ходе иммунного ответа рецепторы распознавания образов (PRR), которые ограничены зародышевой линией, используются для идентификации молекулярных организаций, консервативных среди классов патогенов, таких как вирусная двухцепочечная РНК. Интерфероны (IFN), которые представляют собой секретируемые белки, кодируемые хозяином и подразделяются на три типа (I, II и III), часто участвуют во множественных иммунных взаимодействиях и выполняют как индукцию, так и регуляцию врожденных и адаптивных противовирусных механизмов, когда вирусы инфицируют хозяина. .При возникновении вирусных инфекций экспрессия IFN типа I (как правило, сосредоточена на IFN-α и IFN-β) будет функционировать как основной врожденный ответ противовирусной защиты [1]. Противовирусная активность, проявляемая IFN типа I, напрямую подавляет репликацию вируса. Кроме того, IFN типа I могут опосредовать клеточные иммунные функции как врожденной, так и адаптивной иммунной системы, обеспечивая устойчивость к вирусным инфекциям и поддерживая долгосрочный иммунитет [2]. Из-за очевидных противовирусных функций IFN типа I [3] было проведено множество исследований иммунной активности IFN типа I, и был получен большой объем информации о молекулярных механизмах его биологических действий, иммунной индукции IFN типа I и иммунное уклонение, осуществляемое вирусами [4].Основываясь на многочисленных исследованиях IFN типа I, он используется в качестве иммунного индуктора или лекарственного средства для лечения хронической вирусной инфекции. Среди трех типов IFN, IFN типа III, называемые IFN-λ или IFNL, также играют важную роль в противовирусной иммунной активности [5]. IFN типа III (IFN-λ1, 2 и 3) были обнаружены как интерлейкины (IL) -29, 28a и 28b и обладают многими иммунными активностями, общими с IFN типа I [6, 7]. Позднее был открыт новый член, названный IFN-λ4, который может экспрессироваться только людьми, несущими символ гена (аллель IFNL4-ΔG [rs368234815]) [8].Следует отметить, что врожденные противовирусные ответы на проникновение вирусных частиц были непосредственно опосредованы PRR и не зависели как от путей TLR, так и от RIG-I, а именно, независимо от IFN [9]. Этот вывод иллюстрирует, что ранние события, связанные с врожденной антивирусной активностью, более сложны, чем считалось ранее, и подчеркивает, что исследователям все еще необходимо оценить гораздо больше случаев, чтобы понять ранние этапы борьбы с вирусом-хозяином. В этом обзоре мы пытаемся представить, как были открыты IFN типа III, и обсудить, что было изучено об их роли в посредничестве врожденной / адаптивной иммунной системы и их механизме противовирусной защиты.Наконец, мы предлагаем дальнейшие направления исследований биологии IFN типа III.
Открытие IFN типа III
IFN типа III, включая IFN-λ1, IFN-λ2 и IFN-λ3, были впервые описаны с помощью компьютерных предсказаний в соответствии с данными генома [6, 7]. Открытие IFN-λ4 было сделано путем анализа многих маркеров однонуклеотидного полиморфизма (SNP), расположенных выше хромосомной области IFN-λ3, на основе полногеномных исследований вирусной инфекции гепатита C [8]. Чтобы лучше классифицировать символ гена IFN-λ4, основная информация о местоположении семейства IFN-λ в геноме проиллюстрирована на рис.1. Все четыре члена существуют в области от 19q13.12 до 19q13.13 в длинном (q) плече хромосомы 19. Ген IFNL1 (Il-29) расположен ниже IFNL2 (IL-28a). ), а ген IFNL3 расположен ниже IFNL4 . Три белка (IFN-λ1, IFN-λ2 и IFN-λ3) транскрибируются и транслируются из генов (IFNL1, IFNL2 и IFNL3) и очень похожи друг на друга. В частности, степень сходства между IFN-λ2 и IFN-λ3 составляет примерно 96% на уровне аминокислотной последовательности, а аминокислотная идентичность между IFN-λ1 и IFN-λ2 / IFN-λ3 составляет примерно 81% [6, 7 ].Хотя IFN-λ4 наиболее близок к IFN-λ3, аминокислотная идентичность между IFN-λ3 и IFN-λ4 составляет примерно 30% [10]. Рецептор IFN-λ исследовали посредством сканирования транслированных геномных последовательностей человека на предмет последовательностей, относящихся к рецепторам цитокинов класса II. IFN-λ1-4 рассматриваются как IFN типа III, потому что они передают сигнал через рецепторный комплекс, который отличается от рецептора, используемого IFN типа I и типа II [11, 12]. Следовательно, дальнейшее описание нового типа врожденного противовирусного цитокина вызвало ряд вопросов и потребовало дополнительного понимания роли IFN-λ в противовирусной защите.Здесь мы приводим некоторые новые данные, касающиеся роли IFN-λ в врожденных и адаптивных иммунных ответах, чтобы проиллюстрировать биологическую активность этих IFN и предоставить некоторые справочные предложения для потенциального клинического применения IFN-λ при вирусных инфекциях.
Рис. 1.
Расположение генов семейства IFN-λ в хромосоме 19.
Генерация IFN-λ во время вирусной инфекции
Основным этапом образования IFN является требование обнаружения микробов внутриклеточным методом. рецепторы.Вирусные генетические материалы являются наиболее мощными индукторами IFN-ответов. В цитоплазме 5’-трифосфорилированная или двухцепочечная (ds) РНК распознается геликазами RIG-I и MDA5 домена «DEXD-H box» [13, 14]. В эндосомах ssRNA или dsRNA могут распознаваться Toll-подобными рецепторами (TLR3, TLR7, TLR8 и TLR9) [15-17]. В некоторых случаях ДНК может служить мощным индуктором генерации IFN, но данных о рецепторных системах, опосредующих эти события, не поступало. Сообщалось, что ДНК-зависимый активатор IFN-регуляторных факторов (DAI), также называемый DLM-1 или ZBP1, может распознавать Z- / B-ДНК и запускать экспрессию IFN [18, 19].Взаимодействие между IRF3 и DAI может способствовать ответу IFN на ДНК цитомегаловируса человека [20]; однако DAI, вероятно, увеличивает уровни репликации ВИЧ-1 через путь DAI-NF-κB [21]. Интересно, что из-за перекрестного взаимодействия между системами, чувствительными к ДНК и РНК, РНК-полимераза III распознает некоторые ДНК в качестве матриц и выполняет de novo генерацию 5′-трифосфорилированных РНК с двухцепочечными структурами, которые могут запускать системы IFN через RIG- I путь [22, 23]. Кроме того, были идентифицированы другие новые пути, вовлеченные в систему IFN, запускающую ДНК-сенсор, включая путь cGAS-STING и отсутствующий в меланоме 2 (AIM2) путь инфламмасомы [24–26].В пути cGAS-STING динуклеотид 2 ‘, 3’-GMP-AMP может служить в качестве вторичного эндогенного мессенджера в передаче сигналов врожденного иммунитета, индуцированной чужеродной ДНК, связанной с бактериальной и вирусной инфекцией, и распознаваться рецептором STING, вызывая фосфорилирование TANK-связывающей киназы 1 (TBK1) и IRF3, приводящее к образованию IFN [27]. Для нового рецептора цитоплазматической ДНК AIM2 может формировать инфламмасому с лигандом и ASC (связанный с апоптозом пятнышкообразный белковый домен, содержащий домен активации и рекрутирования каспазы) для активации как каспазы-1, так и NF-κB.Эти сигнальные трансдукции затем запускают систему IFN типа I и пути пироптотической и апоптотической смерти [28-30]. В целом, внутриклеточное восприятие микробов через различные рецепторные системы врожденной иммунной системы запускает передачу сигналов некоторым транскрипционным факторам и, в конечном итоге, приводит к образованию IFN.
Что касается роли факторов транскрипции в экспрессии двух типов IFN, стратегии транскрипции основываются на ядерном факторе (NF) κB и факторах регуляции IFN (IRF).Несмотря на в целом сходную модель транскрипции для экспрессии двух типов, существуют важные различия в механизмах транскрипции, опосредующих экспрессию двух типов [31]. Взяв, например, IFN-α, кластер сайтов связывания IRF существует в промоторах IFN-α, но существуют разные аффинности для связывания IRF3 и IRF7 с соответствующими сайтами, и IRF7 имеет сильную тенденцию связывать IFN-α. промотор для индукции экспрессии гена [32, 33]. Для экспрессии IFN-β как IRF3, так и IRF7 могут хорошо связываться с правильным сайтом в промоторе.Поскольку экспрессия является конститутивной, в то время как экспрессия IRF7 является IFN-специфичной, экспрессия IFN-β является ранним врожденным иммунным ответом, тогда как экспрессия IFN-α является отложенным ответом, но находится на высоком уровне [33, 34]. Что касается экспрессии IFN-λ, то промоторы всех генов IFN-λ имеют сайты связывания для NF-κB и IRF [35, 36]. Следует отметить, что промотор в гене IFN-λ1 имеет высокое сродство к IRF3, тогда как промотор IFN-λ2 / 3 имеет высокое сродство к IRF7. Следовательно, ответы IFN-λ2 / 3 представляют собой замедленную кинетику по сравнению с IFN-λ1 [36].По сравнению с промотором IFN-β человека, кластер дистальных сайтов NF-κB играет важную роль в полной индукции IFN-λ1, и эти сайты активируют промотор IFN-λ1 без IRF-3/7 [37]. Когда путь NF-κB ингибировался в дендритных клетках (DC), генерация IFN-λ серьезно блокировалась, но это ингибирование оказывало незначительное влияние на генерацию IFN типа I [38]. Несмотря на то, что одни и те же транскрипционные факторы участвуют в активации генерации IFN типа I и III, путь NF-κB является ключевым регулятором в генерации IFN-λ, тогда как путь IRFs доминирует над экспрессией IFN типа I.Кроме того, из-за независимого действия NF-κB и IRF-3/7 промоторы IFN-λ кажутся более гибкими, чем промотор IFN-β, в получении отдельных сигналов для активации генерации IFN-λ независимо.
Индукция IFN-λs и сигнальные пути
Все биологические активности цитокинов осуществляются путем взаимодействия со специфическими рецепторами, которые получают стимулы и затем запускают внутриклеточные события через пути передачи сигнала. Стимулы, которые запускают экспрессию генов IFNL , включая вирусы, аналогичны тем трансляциям IFN типа I [6, 7, 39-41].Тем не менее, существуют различия в требованиях к рецепторам и факторам транскрипции между IFN типа I и IFN типа III. Передача сигнала IFN типа I зависит от комплекса IFNAR, который состоит из IFNAR 1 и IFNAR2, в то время как передача сигнала IFN типа III зависит от IFN-λ-специфической цепи IL-28Ra и цепи IL-10R2, которая включает IL-10 и другие члены суперсемейства IL-10 [6, 7]. Когда хозяин обнаруживает ассоциированные с патогенами молекулярные структуры с помощью PRR, могут быть синтезированы IFN типа I и III.Для передачи сигнала, индуцированного IFN типа I, взаимодействия между IFN-α / β и IFNAR запускают активацию специфических рецепторных взаимодействующих тирозинкиназ Jak1 и Tyk2, которые могут фосфорилировать членов сигнального преобразователя и активатора семейства транскрипции (STAT) до запускают димеризацию STAT и активацию активности родственных факторов транскрипции [42]. STAT1 и 2 считаются основными факторами транскрипции, активируемыми IFN, которые вместе с IRF9 образуют тримерный комплекс ISGF3, который управляет транскрипцией стимулированных IFN генов (ISG) (PMID: 7959489).После исследований функций семейства STAT, STAT 3, 4 и 5 также могут быть активированы IFN типа I [43]. Несмотря на различия между рецепторными системами, используемыми IFN типов I и III, внутриклеточные сигнальные программы, активируемые IFN-λ, в некоторой степени схожи. Комбинация IFN-λ и IL-28Rα вызывает конформационное изменение, которое эффективно осуществляет привлечение IL-10R2 в комплекс IFN-λ-IL-28Rα-IL-10R2. Затем активируются связанные с рецептором тирозинкиназы (TYK2 и JAK1), чтобы контролировать фосфорилирование тирозина внутриклеточного домена цепи IL-28Rα [44, 45].Белки STAT отслеживают и связываются с мотивами с фосфотирозином в этом домене и формируют ISGF3, например IFN типа I. ISGF3 перемещается из цитозоля в ядро и связывается с интерферон-стимулированными элементами ответа (ISRE) в промоторах ISG (рис. 2), которые могут продуцировать множество белков с противовирусными функциями [10, 46, 47]. Несмотря на аналогичные модели передачи сигнала, опосредованные IFN типов I и III, IFN типа I могут индуцировать экспрессию ISG с более высокой кинетикой, чем у IFN типа III, а IFN типа III могут продлевать более стабильно высокий уровень экспрессии ISG, чем у типа I. IFNs [46].Остается неясным, обладает ли ИФН типа III иммунной активностью, отличной от иммунной активности ИФН типа I, и обладают ли эти два типа ИФН противовирусной активностью с разной кинетикой.
Рис. 2.
Модель сходства в передаче сигналов, запускаемых рецепторами IFN типов I и III. IFN типов I и III основаны на рецепторных комплексах IFNAR1 / IFNAR2 и IL-28Rα / IL-10R2 соответственно. В некоторой степени внутриклеточные сигнальные трансдукции, индуцированные двумя различными рецепторными комплексами, подобны, особенно с IFN-активированным фактором транскрипции ISGF3, состоящим из SAT1-SAT2-IRF9.ISGF3 может связываться с интерферон-стимулированными элементами ответа (ISRE) в промоторах многочисленных IFN-стимулированных генов (ISG), чтобы запускать экспрессию ISG.
Роль IFN-λ в противовирусных ответах
IFN обычно считаются противовирусными цитокинами при врожденных иммунных ответах. В семействе IFN типа III IFN-λ1, IFN-λ2 и IFN-λ3 представляют собой противовирусную активность против ряда вирусов in vitro [6, 7]. Это быстро вызвало вопросы о функциях IFN-λ в ограничении репликации основных патогенных вирусов человека.Первое сообщение о противовирусном ответе IFN-λ заключается в том, что IFN-λ могут блокировать репликацию вируса гепатита C и вируса гепатита B in vitro [48]. Однако IFN-λ4, который вырабатывается только людьми, несущими аллель IFNL4-ΔG в качестве основного варианта у африканцев и второстепенного варианта у азиатов, связан с неспособностью противостоять инфекции HCV либо спонтанно, либо в ответ на лечение [ 10]. Согласно текущим данным о функциях IFN-λs, многочисленные исследования были сосредоточены на их вкладе в противовирусные иммунные ответы.Рекомбинантные IFN-λ1 и -λ2 ограничивают уровни репликации и цитотоксическую активность вируса простого герпеса (HSV) в клетках HepG2 [40]. После этого IFN-λ1 и -λ2 могут индуцировать экспрессию хемокинов CC, которые способны связываться с входным корецептором ВИЧ-1 CCR5 и ограничивать инфицирование макрофагов ВИЧ-1 [49]. вирусная инфекция, IFN-λ может индуцировать противовирусный фактор Mx1, чтобы ограничить распространение вируса гриппа A в легких; однако IFN-λ не может индуцировать Mx1 для ограничения репликации гепатотропного вируса в печени [50], что позволяет предположить, что IFN-λ играет важную роль во врожденном иммунном ответе в тканях слизистой оболочки.Когда мышей с нокаутированными рецепторами IFN типов I и III заражали вирусом желтой лихорадки (YFV), у этих мышей наблюдались отчетливые изменения в частотах множественных линий иммунных клеток, нарушение активации Т-клеток и серьезное нарушение баланса провоспалительных цитокинов [51]. ]. Кроме того, широкий спектр противовирусной иммунной активности IFN-λ был идентифицирован в печени, легких, головном мозге и кишечном тракте [52–55]. В совокупности репликация функционально и структурно различных вирусов человека нарушается IFN-λ в различных органах и тканях.
Несмотря на сходные противовирусные реакции между IFN типа III и типа I, упомянутые выше, эти два типа IFN существенно различаются в отношении того, на какие клетки они нацелены. Рецепторы (IFNAR) IFN типа I существуют повсеместно, однако рецептор (IL-28Rα) существует только в нескольких типах клеток, включая некоторые классы лейкоцитов, такие как макрофаги, лимфоциты периферической крови, обычные DC, эпителиальные клетки и плазмоцитоидные DC. и поэтому клеточный ответ на IFN-λ ограничен узким спектром типов клеток и тканей [6, 7, 41, 56-60]. Эти данные также указывают на то, что дифференциальная экспрессия рецепторов IFN типа I по сравнению с типом III очевидна. эффекты на биологическую активность этих функционально связанных цитокинов в противовирусном ответе живых организмов.После исследований противовирусных иммунных ответов IFN-λ in vitro рекомбинантный IFN-λ, добавленный экзогенно или экспрессированный из рекомбинантного вируса, был способен ограничивать репликацию вируса у мышей, включая вирус Зика (ZIKV), вирус осповакцины, вирус гриппа А. , вирус гриппа B, коронавирус тяжелого острого респираторного синдрома, метапневмовирус человека, респираторно-синцитиальный вирус, HSV-2 и другие [40, 50, 61-64]. Эти исследования показали, что IFN-λ играет ключевую роль в противовирусных иммунных ответах in vivo и что эпителиальные поверхности являются основным полем битвы за эффективность IFN-λ в врожденных иммунных ответах, которые возникают в дыхательных и желудочно-кишечных трактах.Вышеупомянутые серьезные результаты в основном зависели от модели инфекции (мыши IL-28RA — / — и мыши IFNAR — / -). В литературе сообщается, что мыши STAT1 — / — проявляют более выраженный фенотип по сравнению с мышами IFNAR — / — после вирусного заражения. Это предполагает, что путь STAT играет важную роль во врожденных иммунных ответах на IFN-λ и IFN-γ [64-66].
Несмотря на то, что IFNs служат первой линией иммунной защиты от вторжения вирусных инфекций, вирусы могут применять различные стратегии для подавления противовирусных иммунных ответов IFN.Подобно вирусам, ингибирующим ответы IFN типа I различными методами, как ДНК-, так и РНК-вирусы используют различные стратегии уклонения, чтобы блокировать молекулы, необходимые для экспрессии IFN типа III (например, IRF3), и ингибировать необходимые биологические функции (например, STAT1 / 2). , что приводит к нарушению ответа IFNs типа III [67]. Цитоплазматический белок (E3L) вируса осповакцины может нарушать PKR-зависимый путь для предотвращения противовирусного иммунного ответа, опосредованного IFN-λ. [68] Вирусы Эбола (EBoV) могут генерировать вирусный белок (VP24), который ингибирует ниже по течению от активации IRF3. , блокируя экспрессию IFN-λ [69].Вирусная протеаза (2A pro ) вируса Коксаки может блокировать как путь TLR3, так и пути RIG-I / MDA5 для дальнейшей генерации белков передачи сигнала (TRIF и IPS1) и, следовательно, снижения экспрессии IFN-λ [70]. Несмотря на то, что IFN-λ выполняет важный противовирусный иммунный ответ на норовирусную инфекцию эпителиальных клеток кишечника, вирусный белок (NS1) действует через прямой антагонизм системы IFN-λ и доминирует в тропизме вирусных клеток [71]. Два дополнительных белка ВИЧ-1 (Vpr и Vif) могут связываться с TANK-связывающей киназой 1 (TBK1) и блокировать экспрессию IFN типов I и III в человеческих DC и макрофагах [72], хотя IFN-λ играет ключевую роль в противовирусные иммунные ответы на эпителиальных поверхностях, вирусы изо всех сил стараются уклоняться от противовирусных иммунных ответов для создания инфекции.
Влияние IFN-λ на адаптивные иммунные ответы
Из-за некоторых схожих функций и сигнальных путей между IFN типа I и типа III предполагается, что система IFN-λ обладает некоторыми новыми аспектами врожденной иммунной системы, регулирующей адаптивный иммунитет. отклик. IFN-λ регулирует дифференцировку DC из моноцитов посредством формирования рецепторного комплекса IFN-λ, и стимуляция через этот рецептор специфически индуцирует пролиферацию толерогенных CD4 + CD25 + Foxp3 + регуляторных T (Treg) -клеток, в результате чего в генерации толерогенных ДК, которые могут подавлять функции IFNs [73].Согласно аналогичным результатам исследования рекомбинантный аденовирус, экспрессирующий человеческий IFN-λ1, может снижать секрецию сывороточного IgE и увеличивать количество клеток селезенки CD4 + CD25 + Foxp3 + Treg, ослабляя аллергическое воспаление дыхательных путей [74]. Последующие исследования также отметили, что IFN-λ снижает активность Treg во время развития адаптивного иммунного ответа в более физиологических системах [75]. Таким образом, иммуностимулятор (экстракт выдержанного чеснока) может повышать уровень цитокинов IFN-λ и IL-4 в спленоцитах, стимулированных специфическим опухолевым антигеном, и уменьшать количество Treg-клеток в селезенке [76].Похоже, что IFN-λ позволяет адаптивной иммунной системе снижать иммуносупрессию, регулируемую Treg-клетками. Однако совсем недавно, по сравнению с активностью иммуносупрессии, регулируемой живыми Treg-клетками, мертвые Treg-клетки поддерживают и усиливают супрессорную способность иммунных ответов [77]. Основываясь на этих выводах, мы предполагаем, что дальнейшие исследования роли IFN-λ в снижении активности, индуцированной Treg-клетками, могут пролить свет на то, как контролировать поведение Treg-клеток и повысить эффективность терапии, направленной на контрольные точки рака.
Отдельные подтипы IFN обладают различной эффективностью в избирательной активации подмножеств иммунных клеток для запуска противовирусной иммунной активности, приводящей к выработке устойчивой В- и Т-клеточной памяти [78]. Вскоре после первоначального открытия IFN-λ в некоторых сообщениях было высказано предположение, что IFN-λ может регулировать хелперные Т-клетки и подавлять экспрессию IL-4 и IL-13 в отсутствие ответа IFN-γ [79, 80]. Несмотря на смещение пролиферации Th2 / Th3 в сторону фенотипа Th2, общие однонуклеотидные полиморфизмы в генах IFN-λ и его рецепторных α-субъединиц влияют на передачу сигналов IFN-λ и, таким образом, модулируют баланс Th2 / Th3 и ухудшают терапевтический эффект лечения IFN во время инфекций. [81-83].Вместе с недавним сообщением о том, что IFN-λ блокирует превращение Т-клеток центральной памяти в эффекторные Т-клетки памяти, эти результаты предполагают, что IFN-λ может регулировать наиболее благоприятную среду Т-клеток путем предотвращения дифференцировки Th3 и, следовательно, поддерживать оптимальную адаптивную среду. противовирусный иммунный ответ для борьбы с вирусными инфекциями.
Будущие тенденции применения для клинического лечения
На основании литературы о IFN-λ в настоящее время хорошо установлено, что IFN-λ играет ключевую роль в врожденных / адаптивных иммунных ответах и является лекарством будущего против хронических вирусных инфекций.Эта область срочно требует дальнейшего изучения как базовой биологии, так и терапевтической противовирусной активности IFN-λ. Например, так много исследований улучшили наше понимание эффектов различных подтипов IFN-λ на выведение инфекции HCV [84–89]. Однако остается путаница в отношении влияния подтипов IFN-λ на иммунный ответ на вирусные инфекции. Несмотря на то, что инфицированные HCV пациенты с аллелем IFNL4-ΔG, как правило, не избавляются от инфекции HCV, а IFN-λ4 секретируется лишь незначительно, эти пациенты имеют более низкую вирусную нагрузку HCV без лечения [10].Совсем недавно было сообщено, что генетические варианты в IFNL4 обладают различной эффективностью в устранении инфекции HCV в китайской популяции хань [90]. В частности, остается неясным, существует ли взаимосвязь между противовирусными иммунными ответами моделей мышей с инвазивной инфекцией ВГС и отсутствием цитокинов (IFN-λ1 или IFN-λ4) [91]. Такие знания могут выявить новые пути улучшения подтипов IFN-λ при разработке вакцин против ВГС. Из-за частично перекрывающихся сигнальных путей (RIG-I, MDA5 и MAVS), опосредованных IFN типа I и типа III, парадоксальная иммунная активность может осуществляться двумя типами IFN.Независимо от подтипов IFN, RIG-I активирует две различные категории ISG, одну JAK-STAT-зависимую, а другую JAK-STAT-независимую, которые согласованно вносят вклад в противовирусный иммунный ответ на инфекцию HEV [92]. Однако стойкая активация JAK-STAT-зависимого сигнального пути позволяет HEV-инфицированным клеткам противостоять экзогенному воздействию IFN, в то время как истощение рецепторов IFN-λ или MAVS (митохондриальный антивирусный сигнальный белок) приводит к противовирусному иммунному ответу, индуцированному IFN, предполагая, что постоянное присутствие IFN-λ способствует возникновению инфекции HEV [93].Вместе с недавним сообщением о противовирусной иммунной активности, опосредованной IFN-λ, нам все еще не хватает важной информации об основных функциях IFN-λ. Какова молекулярная природа взаимодействий между цитокином и рецептором? Совсем недавно кристаллизованный тройной комплекс (комплекс IFN-λ-IL28Ra-IL-10R2) подчеркивает пластичность передачи сигналов IFN-λ и его терапевтический потенциал [94]. Лучшее понимание взаимодействия между IFN-λ и его рецепторами может пролить свет на то, что активирует передачу сигналов, а также может способствовать развитию цитокинов с измененной функцией.Что касается передачи сигналов, опосредованной IFN-λ, то в настоящее время известно, что IFN I и III типов индуцируют сходные пути передачи сигналов. Несмотря на то, что JAK-STAT-зависимая передача сигнала осуществляется IFN как I, так и III типа, у нас все еще очень ограниченные знания о других активируемых IFN-λ путях, которые могут потенциально влиять на иммунную активность IFN-λ.
Для связи между врожденным иммунным ответом и IFN-λ необходимы дальнейшие исследования относительного вклада полиморфизмов IFNL в иммунную защиту хозяина.Например, недавно сообщалось, что полиморфизмы IFNL3 и IRF7 могут модулировать иммунный ответ против HSV-1 при болезни Альцгеймера [95] и что полиморфизм IFNL3 также играет роль в иммунном ответе на терапию IFN у пациентов с хроническим HBV и HCV [95] 96, 97], предполагая, что генетический полиморфизм IFNL3 может играть очевидную роль во врожденной защите. Что касается роли IFN-λ в адаптивном иммунном ответе, нам необходимо определить, какие клетки адаптивной иммунной системы отвечают на IFN-λ, и роль эндогенного IFN-λ в поддержании и развитии оптимальных адаптивных иммунных ответов на противостоять вирусной инфекции.Связанные с этим исследования могут внести вклад в разработку терапевтических препаратов IFN-λ и адъювантов для вакцин, связанных с IFN-λ.
Заключение
В заключение, IFN типа III были идентифицированы как новый класс цитокинов, которые представляют собой специализированные вирус-индуцированные IFN с иммунными и биологическими функциями, как перекрывающимися с IFN типа I, так и отличными от них. Лучшее понимание связанных функций и взаимодействий между различными противовирусными системами в иммунной системе может помочь исследователям в разработке терапевтических методов или иммунных регуляторов, включающих IFN-λ, для вторжения вирусных патогенов в организме хозяина и установления долговременного иммунитета без чрезмерного воздействия. активация воспаления.
Благодарности
Работа поддержана Национальным фондом естественных наук Китая (№ 31302100; 31700763; 81760287).
Заявление о раскрытии информации
Все авторы заявляют, что у них нет конкурирующих интересов.
Список литературы
- Гарсия-Састре А., Бирон, Калифорния: Интерфероны типа 1 и отношения вирус-хозяин: урок разрядки.Наука 2006; 312: 879-882.
- Мальмгаард Л. Индукция и регулирование интерферонов при вирусных инфекциях. J Interferon Cytokine Res 2004; 24: 439-454.
- Айзекс А., Линденманн Дж .: Вмешательство вирусов.I. Интерферон. Авторы A. Isaacs и J. Lindenmann, 1957. J. Interferon Res 1987; 7: 429-438.
- Самуэль CE: Противовирусное действие интерферонов. Clin Microbiol Rev 2001; 14: 778-809.
- Hamana A, Takahashi Y, Uchida T, Nishikawa M, Imamura M, Chayama K, Takakura Y: Оценка противовирусного эффекта интерферонов I, II и III типа на противовирусный устойчивый вирус гепатита C прямого действия.Antiviral Res 2017; 146: 130-138.
- Котенко С.В., Галлахер Г., Баурин В.В., Льюис-Антес А., Шен М., Шах Н.К., Лангер Дж. А., Шейх Ф., Диккеншитс Х., Доннелли Р.П.: IFN-лямбды опосредуют противовирусную защиту через отдельный рецепторный комплекс цитокинов класса II. Nat Immunol 2003; 4: 69-77.
- Шеппард П., Киндсфогель В., Сюй В., Хендерсон К., Шлуцмайер С., Уитмор Т. Е., Куэстнер Р., Гарригес Ю., Биркс С., Рорабак Дж., Острандер С., Донг Д., Шин Дж., Преснелл С., Фокс Б., Холдеман Б., Купер И. , Taft D, Gilbert t, Grant FJ et al .: IL-28, IL-29 и их рецептор цитокинов класса II IL-28R.Nat Immunol 2003; 4: 63-68.
- Прокунина-Олссон Л., Мучмор Б., Тан В., Пфайффер Р. М., Парк Х, Диккеншитс Х, Херготт Д., Портер-Гилл П., Муми А., Кохар I, Чен С., Бренд N, Тарвей М., Лю Л., Шейх Ф, Астемборски J, Бонковский HL, Эдлин BR, Howell CD, Morgan TR и др.: Вариант выше IFNL3 (IL28B), создающий новый ген интерферона IFNL4, связан с нарушением клиренса вируса гепатита C.Nat Genet 2013; 45: 164-171.
- Паладино П., Каммингс Д.Т., Нойс Р.С., Моссман К.Л.: IFN-независимый ответ на проникновение вирусных частиц обеспечивает первую линию противовирусной защиты, которая не зависит от TLR и гена, индуцируемого ретиноевой кислотой I. J Immunol 2006; 177: 8008-8016.
- О’Брайен Т.Р., Прокунина-Олссон Л., Доннелли Р.П.: IFN-lambda4: парадоксальный новый член семейства интерфероновых лямбда.J Interferon Cytokine Res 2014; 34: 829-838.
- de Weerd NA, Nguyen T: Интерфероны и их рецепторы — распределение и регуляция. Immunol Cell Biol 2012; 90: 483-491.
- Hamming OJ, Terczynska-Dyla E, Vieyres G, Dijkman R, Jorgensen SE, Akhtar H, Siupka P, Pietschmann T., Thiel V, Hartmann R: сигналы интерферона лямбда 4 через рецептор IFNlambda для регулирования противовирусной активности против HCV и коронавирусов.EMBO J 2013; 32: 3055-3065.
- Като Х, Такеучи О, Сато С., Ёнеяма М., Ямамото М., Мацуи К., Уэмацу С., Юнг А., Кавай Т., Исии К.Дж., Ямагути О, Оцу К., Цудзимура Т., Кох К.С., Рейс и Соуза К., Мацуура Ю., Fujita T, Akira S: Различная роль геликаз MDA5 и RIG-I в распознавании РНК-вирусов.Природа 2006; 441: 101-105.
- Йонеяма М., Кикучи М., Нацукава Т., Синобу Н., Имаидзуми Т., Миягиши М., Тайра К., Акира С., Фудзита Т.: РНК-геликаза RIG-I играет важную роль в индуцированных двухцепочечной РНК врожденных противовирусных реакциях. Нат Иммунол 2004; 5: 730-737.
- Alexopoulou L, Holt AC, Medzhitov R, Flavell RA: Распознавание двухцепочечной РНК и активация NF-kappaB с помощью Toll-подобного рецептора 3.Nature 2001; 413: 732-738.
- Diebold SS, Kaisho T, Hemmi H, Akira S, Reis e Sousa C: врожденные противовирусные реакции посредством TLR7-опосредованного распознавания одноцепочечной РНК. Наука 2004; 303: 1529-1531.
- Heil F, Hemmi H, Hochrein H, Ampenberger F, Kirschning C, Akira S, Lipford G, Wagner H, Bauer S: Видоспецифическое распознавание одноцепочечной РНК через толл-подобные рецепторы 7 и 8.Наука 2004; 303: 1526-1529.
- Ким К., Хайрутдинов Б.И., Ли С.К., Чеонг Х.К., Кан С.В., Пак Х., Ли С., Ким Ю.Г., Джи Дж., Рич А., Ким К.К., Чон Й.Х .: Структура раствора домена Zbeta ДНК-зависимого активатора IFN человека. -регуляторные факторы и способы их связывания с B- и Z-ДНК.Proc Natl Acad Sci USA 2011; 108: 6921-6926.
- Takaoka A, Wang Z, Choi MK, Yanai H, Negishi H, Ban T, Lu Y, Miyagishi M, Kodama T, Honda K, Ohba Y, Taniguchi T: DAI (DLM-1 / ZBP1) — датчик цитозольной ДНК и активатор врожденного иммунного ответа. Природа 2007; 448: 501-505.
- ДеФилиппис В.Р., Альварадо Д., Сали Т., Ротенбург С., Фру К. Цитомегаловирус человека вызывает интерфероновый ответ через датчик ДНК ZBP1. J Virol 2010; 84: 585-598.
- Hayashi T, Nishitsuji H, Takamori A, Hasegawa A, Masuda T., Kannagi M: ДНК-зависимый активатор IFN-регуляторных факторов усиливает транскрипцию ВИЧ-1 через NF-kappaB.Microbes Infect 2010; 12: 937-947.
- Ablasser A, Bauernfeind F, Hartmann G, Latz E, Fitzgerald KA, Hornung V: RIG-I-зависимое восприятие поли (dA: dT) посредством индукции РНК-полимеразы III-транскрибируемого промежуточного звена РНК. Нат Иммунол 2009; 10: 1065-1072.
- Chiu YH, Macmillan JB, Chen ZJ: РНК-полимераза III обнаруживает цитозольную ДНК и индуцирует интерфероны типа I через путь RIG-I.Cell 2009; 138: 576-591.
- Fernandes-Alnemri T, Yu JW, Datta P, Wu J, Alnemri ES: AIM2 активирует инфламмасомы и гибель клеток в ответ на цитоплазматическую ДНК. Природа 2009; 458: 509-513.
- Hornung V, Ablasser A, Charrel-Dennis M, Bauernfeind F, Horvath G, Caffrey DR, Latz E, Fitzgerald KA: AIM2 распознает цитозольную дцДНК и с ASC формирует воспаление, активирующее каспазу-1.Природа 2009; 458: 514-518.
- Sun L, Wu J, Du F, Chen X, Chen ZJ: Циклическая GMP-AMP-синтаза — это цитозольный ДНК-сенсор, который активирует путь интерферона типа I. Наука 2013; 339: 786-791.
- Като К., Омура Х, Ишитани Р., Нуреки О: Циклический GMP-AMP как эндогенный второй мессенджер в врожденной иммунной передаче сигналов цитозольной ДНК.Annu Rev Biochem 2017; 86: 541-566.
- Compan V, Martin-Sanchez F, Baroja-Mazo A, Lopez-Castejon G, Gomez AI, Verkhratsky A, Brough D, Pelegrin P: связанный с апоптозом пятнышкообразный белок, содержащий CARD, образует пятнышки, но не активирует каспазу-1 в отсутствие NLRP3 при набухании макрофагов.J Immunol 2015; 194: 1261-1273.
- Sagulenko V, Thygesen SJ, Sester DP, Idris A, Cridland JA, Vajjhala PR, Roberts TL, Schroder K, Vince JE, Hill JM, Silke J, Stacey KJ: инфламмасомы AIM2 и NLRP3 активируют пути апоптотической и пироптотической смерти через ASC. Cell Death Differ 2013; 20: 1149-1160.
- Унтерхольцнер Л: Ответ интерферона на внутриклеточную ДНК: почему так много рецепторов? Иммунобиология 2013; 218: 1312-1321.
- Свецки М., Колонна М.: Интерфероны типа I: разнообразие источников, пути продуцирования и влияние на иммунные ответы.Curr Opin Virol 2011; 1: 463-475.
- Genin P, Vaccaro A, Civas A: роль дифференциальной экспрессии человеческого интерферона-генов в противовирусном иммунитете. Cytokine Growth Factor Rev 2009; 20: 283-295.
- Хонда К., Янаи Х, Негиси Х, Асагири М., Сато М., Мизутани Т., Шимада Н., Охба Й, Такаока А, Йошида Н., Танигути Т.: IRF-7 является главным регулятором интерферон-зависимых иммунных ответов типа I.Природа 2005; 434: 772-777.
- Мари I, Дурбин JE, Леви DE: Дифференциальная вирусная индукция отдельных генов интерферона-альфа посредством положительной обратной связи через фактор регуляции интерферона-7. EMBO J 1998; 17: 6660-6669.
- Оногучи К., Йонеяма М., Такемура А., Акира С., Танигучи Т., Намики Х., Фудзита Т.: Вирусные инфекции активируют гены интерферона I и III типов посредством общего механизма.J Biol Chem 2007; 282: 7576-7581.
- Osterlund PI, Pietila TE, Veckman V, Kotenko SV, Julkunen I. Члены семейства регуляторных факторов IFN по-разному регулируют экспрессию генов IFN типа III (IFN-лямбда). J Immunol 2007; 179: 3434-3442.
- Томсон С.Дж., Го Ф.Г., Бэнкс Х., Краусгрубер Т., Котенко С.В., Фоксвелл Б.М., Удалова И.А.: Роль мобильных элементов в регуляции экспрессии гена IFN-lambda1.Proc Natl Acad Sci USA 2009; 106: 11564-11569.
- Iversen MB, Ank N, Melchjorsen J, Paludan SR: Экспрессия интерферона типа III (IFN) в слизистой оболочке влагалища опосредуется в основном дендритными клетками и проявляет более сильную зависимость от NF-kappaB, чем IFN типа I. J Virol 2010; 84: 4579-4586.
- Ank N, Iversen MB, Bartholdy C, Staeheli P, Hartmann R, Jensen UB, Dagnaes-Hansen F, Thomsen AR, Chen Z, Haugen H, Klucher K, Paludan SR: важная роль интерферона III типа (IFN-lambda / IL-28) с TLR-индуцированной противовирусной активностью. J Immunol 2008; 180: 2474-2485.
- Ank N, West H, Bartholdy C, Eriksson K, Thomsen AR, Paludan SR: Лямбда-интерферон (IFN-lambda), IFN типа III, индуцируется вирусами и IFN и проявляет мощную противовирусную активность против некоторых вирусных инфекций in vivo.J Virol 2006; 80: 4501-4509.
- Дурбин Р.К., Котенко С.В., Дурбин Ю.Е.: Индукция и функция интерферона на поверхности слизистой оболочки. Immunol Rev 2013; 255: 25-39.
- Старк Г. Р., Керр И. М., Уильямс Б. Р., Сильверман Р. Х., Шрайбер Р. Д.: Как клетки реагируют на интерфероны.Annu Rev Biochem 1998; 67: 227-264.
- Ank N, West H, Paludan SR: IFN-lambda: новые противовирусные цитокины. J Interferon Cytokine Res 2006; 26: 373-379.
- Gad HH, Dellgren C, Hamming OJ, Vends S, Paludan SR, Hartmann R: Интерферон-лямбда функционально является интерфероном, но структурно связан с семейством интерлейкинов-10.J Biol Chem 2009; 284: 20869-20875.
- Микнис З.Дж., Маграчева Э., Ли В., Зданов А., Котенко С.В., Влодавер А.: Кристаллическая структура человеческого интерферона-лямбда1 в комплексе с его высокоаффинным рецептором интерферон-лямбдаR1. J Mol Biol 2010; 404: 650-664.
- Марчелло Т., Гракуи А., Барба-Спаэт Г., Махлин Е.С., Котенко С.В., Макдональд М.Р., Райс К.М.: Интерфероны альфа и лямбда ингибируют репликацию вируса гепатита С с помощью четкой передачи сигнала и кинетики регуляции генов.Гастроэнтерология 2006; 131: 1887-1898.
- Zhou Z, Hamming OJ, Ank N, Paludan SR, Nielsen AL, Hartmann R: Интерферон III типа (IFN) индуцирует IFN-подобный ответ I типа в ограниченном подмножестве клеток через сигнальные пути, включающие как путь Jak-STAT, так и митоген-активируемые протеинкиназы.J Virol 2007; 81: 7749-7758.
- Робек М.Д., Бойд Б.С., Чисари Ф.В.: Лямбда-интерферон подавляет репликацию вирусов гепатита В и С. J Virol 2005; 79: 3851-3854.
- Хоу В., Ван X, Е Л., Чжоу Л., Ян З. К., Ридель Э, Хо В. З.: Лямбда-интерферон подавляет инфицирование макрофагов вирусом иммунодефицита человека 1 типа.J Virol 2009; 83: 3834-3842.
- Mordstein M, Kochs G, Dumoutier L, Renauld JC, Paludan SR, Klucher K, Staeheli P: Интерферон-лямбда способствует врожденному иммунитету мышей против вируса гриппа A, но не против гепатотропных вирусов. PLoS Pathog 2008; 4: e1000151.
- Дуам Ф., Сото Альбрехт Ю.Е., Хребикова Г., Садимин Е., Дэвидсон С., Котенко С.В., Плосс А: Интерферон-опосредованная передача сигналов типа III имеет решающее значение для борьбы с живой ослабленной вирусной инфекцией желтой лихорадки in vivo.MBio 2017; 8: e00819-17.
- Boisvert M, Shoukry NH: Интерфероны типа III при вирусной инфекции гепатита С. Фронт Иммунол 2016; 7: 628.
- Davidson S, McCabe TM, Crotta S, Gad HH, Hessel EM, Beinke S, Hartmann R, Wack A: IFNlambda является мощным противогриппозным терапевтическим средством без побочных воспалительных эффектов лечения IFNalpha.EMBO Mol Med 2016; 8: 1099-1112.
- Lazear HM, Daniels BP, Pinto AK, Huang AC, Vick SC, Doyle SE, Gale M., Jr Klein RS, Diamond MS: Интерферон-лямбда ограничивает нейроинвазию вируса Западного Нила, ужесточая гематоэнцефалический барьер. Sci Transl Med 2015; 7: 284ra259.
- Ли С., Болдридж MT: Интерферон-лямбда: мощный регулятор кишечных вирусных инфекций.Фронт Иммунол 2017; 8: 749.
- Lalle E, Bordi L, Caglioti C, Garbuglia AR, Castilletti C, Taibi C, Cristofari F, Capobianchi MR: повышенная регуляция IFN-альфа-рецептора-1 в PBMC от пациентов, ранее не инфицированных HCV, несущих генотип cc. возможная роль IFN-лямбда. PLoS One 2014; 9: e93434.
- Lazear HM, Nice TJ, Diamond MS: Интерферон-лямбда: иммунные функции на барьерных поверхностях и за их пределами.Иммунитет 2015; 43: 15-28.
- Sommereyns C, Paul S, Staeheli P, Michiels T: IFN-lambda (IFN-lambda) экспрессируется тканезависимым образом и в первую очередь действует на эпителиальные клетки in vivo. PLoS Pathog 2008; 4: e1000017.
- Stiff A, Carson W: Исследования интерферона-лямбда для лечения рака.J Innate Immun 2015; 7: 243-250.
- Йошио С., Канто Т., Курода С., Мацубара Т., Хигаситани К., Какита Н., Исида Х., Хирамацу Н., Нагано Х, Сугияма М., Мурата К., Фукухара Т., Мацуура Ю., Хаяси Н., Мизоками М., Такехара Т.: Человеческая кровь дендритные клетки антиген 3 (BDCA3) (+) дендритные клетки являются мощным продуцентом интерферона-лямбда в ответ на вирус гепатита С.Гепатология 2013; 57: 1705-1715.
- Бартлетт Н.В., Буттигег К., Котенко С.В., Смит Г.Л.: Лямбды мышиного интерферона (интерфероны типа III) проявляют мощную противовирусную активность in vivo на модели поксвирусной инфекции. J Gen Virol 2005; 86: 1589-1596.
- Jagger BW, Miner JJ, Cao B, Arora N, Smith AM, Kovacs A, Mysorekar IU, Coyne CB, Diamond MS: стадия беременности и передача сигналов IFN-лямбда регулируют инфекцию ZIKV в утробе матери.Cell Host Microbe 2017; 22: 366-376.
- Kim BJ, Cho SW, Jeon YJ, An S, Jo A, Lim JH, Kim DY, Won TB, Han DH, Rhee CS, Kim HJ: Интраназальная доставка ДНК Duox2 с использованием катионного полимера может предотвратить острую вирусную инфекцию гриппа A in vivo легкое. Appl Microbiol Biotechnol. 2018; 102: 105-115.
- Mordstein M, Neugebauer E, Ditt V, Jessen B, Rieger T, Falcone V, Sorgeloos F, Ehl S, Mayer D, Kochs G, Schwemmle M, Gunther S, Drosten C, Michiels T, Staeheli P: интерферон лямбда визуализирует эпителиальные клетки респираторного и желудочно-кишечного трактов, устойчивых к вирусным инфекциям.J Virol 2010; 84: 5670-5677.
- Johnson TR, Mertz SE, Gitiban N, Hammond S, Legallo R, Durbin RK, Durbin JE: Роль врожденных IFNs в определении иммунопатологии респираторно-синцитиального вируса. J Immunol 2005; 174: 7234-7241.
- Karst SM, Wobus CE, Lay M, Davidson J, Virgin HWt: STAT1-зависимый врожденный иммунитет к норволк-подобному вирусу.Наука 2003; 299: 1575–1578.
- Katze MG, He Y, Gale M, Jr: Вирусы и интерферон: борьба за превосходство. Nat Rev Immunol 2002; 2: 675-687.
- Bandi P, Pagliaccetti NE, Robek MD: Ингибирование активности интерферона типа III иммуномодулирующими белками ортопоксвируса.J Interferon Cytokine Res 2010; 30: 123-134.
- He F, Melen K, Maljanen S, Lundberg R, Jiang M, Osterlund P, Kakkola L, Julkunen I. Белок VP24 эболавируса вмешивается в врожденные иммунные ответы, подавляя экспрессию гена интерферона-лямбда1. Вирусология 2017; 509: 23-34.
- Линд К., Сведин Е., Домсген Е., Капелл С., Лайтинен О, Молл М., Флодстром-Тулберг М.: Вирус Коксаки противостоит врожденному иммунному ответу хозяина, блокируя экспрессию интерферона III типа. J Gen Virol 2016; 97: 1-12.
- Lee S, Wilen CB, Orvedahl A, McCune BT, Kim KW, Orchard RC, Peterson ST, Nice TJ, Baldridge MT, Virgin HW: Тропизм клеток норовируса определяется комбинаторным действием вирусного неструктурного белка и цитокина хозяина.Cell Host Microbe 2017; 22: 449-459.
- Harman AN, Nasr N, Feetham A, Galoyan A, Alshehri AA, Rambukwelle D, Botting RA, Hiener BM, Diefenbach E, Diefenbach RJ, Kim M, Mansell A, Cunningham AL: ВИЧ блокирует индукцию интерферона в дендритных клетках человека и макрофагах Нарушение регуляции TBK1.J Virol 2015; 89: 6575-6584.
- Mennechet FJ, Uze G: Обработанные интерфероном лямбда дендритные клетки специфически индуцируют пролиферацию FOXP3-экспрессирующих супрессорных Т-клеток. Кровь 2006; 107: 4417-4423.
- Li Y, Gao Q, Yuan X, Zhou M, Peng X, Liu X, Zheng X, Xu D, Li M: аденовирус, экспрессирующий IFN-lambda1 (IL-29), ослабляет аллергическое воспаление дыхательных путей и гиперреактивность дыхательных путей при экспериментальной астме.Инт Иммунофармакол 2014; 21: 156-162.
- Morrow MP, Pankhong P, Laddy DJ, Schoenly KA, Yan J, Cisper N, Weiner DB: сравнительная способность IL-12 и IL-28B регулировать популяции Treg и повышать адаптивный клеточный иммунитет. Кровь 2009; 113: 5868-5877.
- Larypoor M, Bayat M, Zuhair MH, Akhavan Sepahy A, Amanlou M: Оценка количества Treg-клеток CD4 (+) CD25 (+) FoxP3 (+) у нормальных мышей, подвергшихся действию AFB1 и обработанных экстрактом старого чеснока.Cell J 2013; 15: 37-44.
- Maj T, Wang W, Crespo J, Zhang H, Wei S, Zhao L, Vatan L, Shao I, Szeliga W, Lyssiotis C, Liu JR, Kryczek I, Zou W: Окислительный стресс контролирует апоптоз регуляторных Т-клеток и активность супрессоров и Устойчивость к PD-L1-блокаде в опухоли. Nat Immunol.2017; 18: 1332.
- Берри CM: Понимание терапии подтипа интерферона при вирусных инфекциях: использование силы врожденной иммунной системы. Cytokine Growth Factor Rev 2016; 31: 83-90.
- Dai J, Megjugorac NJ, Gallagher GE, Yu RY, Gallagher G: IFN-lambda1 (IL-29) ингибирует экспрессию GATA3 и подавляет ответы Th3 в человеческих наивных Т-клетках и Т-клетках памяти.Кровь 2009; 113: 5829-5838.
- Джордан В.Дж., Эскдейл Дж., Сринивас С., Пекарек В., Келнер Д., Родия М., Галлахер Г.: Человеческий интерферон лямбда-1 (IFN-lambda1 / IL-29) модулирует ответ Th2 / Th3. Genes Immun 2007; 8: 254-261.
- Бхушан А., Гош С., Бхаттачарджи С., Чиннасвами С. Смешение с помощью однонуклеотидного полиморфизма rs117648444 (P70S) влияет на ассоциацию вариантов локуса интерферона лямбда с ответом на терапию интерферон-альфа-рибавирином у пациентов с вирусным гепатитом с хроническим генотипом инфекции.J Interferon Cytokine Res 2017; 37: 369-382.
- Эгли А., Сантер Д.М., О’Ши Д., Тиррелл Д.Л., Хоутон М.: Влияние семейства интерферонов-лямбда на врожденный и адаптивный иммунный ответ на вирусные инфекции. Emerg Microbes Infect 2014; 3: e51.
- Hegazy AN, Peine M, Helmstetter C, Panse I, Frohlich A, Bergthaler A, Flatz L, Pinschewer DD, Radbruch A, Lohning M: Интерфероны направляют перепрограммирование клеток Th3 для создания стабильного GATA-3 (+) T-бет (+ ) подмножество клеток с комбинированными функциями клеток Th3 и Th2.Иммунитет 2010; 32: 116-128.
- Griffiths SJ, Dunnigan CM, Russell CD, Haas JG: Роль полиморфизмов интерферон-лямбда-локуса в гепатите C и других инфекционных заболеваниях. J Innate Immun 2015; 7: 231-242.
- Hermant P, Michiels T: Интерферон-лямбда в контексте вирусных инфекций: продукция, ответ и терапевтические последствия.J Innate Immun 2014; 6: 563-574.
- Лейдлоу С.М., Дастин Л.Б.: Интерферон-лямбда: возможности, риски и неопределенности в борьбе с ВГС. Фронт Иммунол 2014; 5: 545.
- Nguyen LT, Van Nguyen D, Carr MJ, Hall WW, Nguyen LA: Ассоциация полиморфизмов лямбда-интерферона с повышенными исходными вирусными нагрузками при хронической инфекции вируса гепатита C генотипа 6.Arch Virol. 2018; 163: 115-124.
- Сахаи Ф., Газанфари М., Вазири Ф., Джамнани Ф.Р., Давари М., Гарибзаде С., Фатех Р., Абдолрахими Ф., Дизаджи С.П., Фатех А., Сиадат С.Д .: Влияние генетической изменчивости генов IL28B, IFNL4 и HLA на реакцию на лечение хронических заболеваний. инфекция вирусом гепатита С.Infect Genet Evol 2017; 54: 330-337.
- Wieczorek-Godlewska R, Ploski R, Perkowska-Ptasinska A, Tronina O, Sadowska A, Pacholczyk M, Lisik W., Durlik M: Влияние полиморфизма интерферона лямбда-3 реципиента и донора на течение инфекции ВГС после трансплантации печени.Clin Exp Hepatol 2017; 3: 152-158.
- Хуанг П, Яо Й, Юэ М., Тиан Т, Чен Х, Чен М, Ван Дж, Чжан И, Ю Р: Генетические варианты интерферона-лямбда 4 влияют на клиренс ВГС в китайской ханьской популяции. Sci Rep 2017; 7: 42408.
- Ласфар А., Злоза А., Коэн-Солал К.А.: IFN-лямбда-терапия: текущее состояние и перспективы на будущее.Drug Discov Today 2016; 21: 167-171.
- Xu L, Wang W, Li Y, Zhou X, Yin Y, Wang Y, de Man RA, van der Laan LJW, Huang F, Kamar N, Peppelenbosch MP, Pan Q: RIG-I — ключевой антивирусный ген, стимулируемый интерфероном. против вируса гепатита Е независимо от выработки интерферона. Гепатология 2017; 65: 1823-1839.
- Инь X, Ли X, Амбардекар С., Ху З., Ломм С., Фэн З .: Вирус гепатита Е сохраняется в присутствии интерферонового ответа типа III. PLoS Pathog 2017; 13: e1006417.
- Mendoza JL, Schneider WM, Hoffmann HH, Vercauteren K, Jude KM, Xiong A, Moraga I, Horton TM, Glenn JS, de Jong YP, Rice CM, Garcia KC: комплекс IFN-lambda-IFN-lambdaR1-IL-10Rbeta Выявлены структурные особенности, лежащие в основе функциональной пластичности IFN типа III.Иммунитет 2017; 46: 379-392.
- Costa AS, Agostini S, Guerini FR, Mancuso R, Zanzottera M, Ripamonti E, Racca V, Nemni R, Clerici M: Модуляция иммунных ответов на вирус простого герпеса типа 1 с помощью полиморфизмов IFNL3 и IRF7: исследование болезни Альцгеймера. J Alzheimers Dis 2017; 60: 1055-1063.
- Индольфи Г., Аззари С., Рести М.: Полиморфизмы в гене IFNL3 / IL28B и гепатит С: от взрослых к детям. World J Gastroenterol 2014; 20: 9245-9252.
- Lin Z, Zhang J, Ma X, Yang S, Tian N, Lin X, Zhou S, Liu L, Gao Y: Роль генетических полиморфизмов интерферона лямбда-3 в ответ на терапию интерфероном у пациентов с хроническим гепатитом B: обновленная версия Meta Анализ.Hepat Mon 2016; 16: e37534.
Автор Контакты
Юнхао Ху и Синь Цао
Колледж ветеринарной медицины,
Сельскохозяйственный университет Ганьсу Ланьчжоу, 730000, Ганьсу (Китай)
Электронная почта [email protected]; [email protected]
Подробности статьи / публикации
Предварительный просмотр первой страницы
Поступило: 14 апреля 2018 г.
Принято: 7 ноября 2018 г.
Опубликовано онлайн: 15 ноября 2018 г.
Дата выпуска: ноябрь 2018 г.
Количество страниц для печати: 13
Количество рисунков: 2
Количество столов: 0
ISSN: 1015-8987 (печатный)
eISSN: 1421-9778 (онлайн)
Для дополнительной информации: https: // www.karger.com/CPB
Лицензия открытого доступа / Дозировка лекарства / Заявление об ограничении ответственности
Эта статья находится под международной лицензией Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 (CC BY-NC-ND). Использование и распространение в коммерческих целях, а также любое распространение измененных материалов требует письменного разрешения. Дозировка лекарств: авторы и издатель приложили все усилия, чтобы гарантировать, что выбор и дозировка лекарств, указанные в этом тексте, соответствуют текущим рекомендациям и практике на момент публикации.Тем не менее, ввиду продолжающихся исследований, изменений в правительственных постановлениях и постоянного потока информации, касающейся лекарственной терапии и реакций на них, читателю настоятельно рекомендуется проверять листок-вкладыш для каждого препарата на предмет любых изменений показаний и дозировки, а также дополнительных предупреждений. и меры предосторожности. Это особенно важно, когда рекомендованным агентом является новое и / или редко применяемое лекарство. Отказ от ответственности: утверждения, мнения и данные, содержащиеся в этой публикации, принадлежат исключительно отдельным авторам и соавторам, а не издателям и редакторам.Появление в публикации рекламы и / или ссылок на продукты не является гарантией, одобрением или одобрением рекламируемых продуктов или услуг или их эффективности, качества или безопасности. Издатель и редактор (-ы) не несут ответственности за любой ущерб, причиненный людям или имуществу в результате любых идей, методов, инструкций или продуктов, упомянутых в контенте или рекламе.
Стандартизованный экстракт эхинацеи пурпурной стимулирует IFN-специфический ответ врожденного иммунного моноцита через JAK1-STAT1-зависимую экспрессию гена и эпигенетический контроль эндогенных ретровирусных последовательностей
3. Manayi, A., M.Vazirian, andS.Saeidnia, Echinaceapurpurea: «Фармакология,« фитохимия »и
методов анализа. 17): .63-72.
4. Karsch-Volk, M., B.Barrett, andK.Linde, Echinaceaforpreventandleatingthecreams.JAMA,
2015 .313 (6): p.618-9.
5. Binns, SE, etal., Фитохимическая разновидностьinechinacea из корнейицветков диких и
культурных популяций. JAgricFood Chem, 2002.50 (13): p.3673-87.
6. Schapowal, A., P.Klein, иSLJohnston, Echinacea снижает риск повторного респираторного лечения
инфекций и осложнений: a Метаанализрандомизированныхконтролируемыхиспытаний.AdvTher, 2015.32 (3): p.
187-200.
7. Hudson, JB, ApplicationsofthephytomedicineEchinaceapurpurea (PurpleConeflower) ininfectious
disease.JBiomedBiotechnol, 2012.2012: стр. 769896.
8. Chicca, A., Etal., Синергетические иммуномофармакологические эффектыN-алкиламидов в Echinaceapurpurea
травяных экстрактов.IntImmunopharmacol, 2009.9 (7-8): p.850-8.
9. Dalby-Brown, L., Etal., Синергетическое антиоксидантное действие алкамидов, производных кофейной кислоты и
полисахаридных фракций из эхинацеи пурпурной. Онвитроокисление липопротеинов низкой плотности человека
. JAgricFoodChem, 2005.53 (24): p.9413-23.
10. Benson, JM, etal., Echinaceapurpureaextractsmodulatemurinedendriticcellfateand function.Food
ChemToxicol, 2010.48 (5): стр.№1170-7.
11. Ричи, М.Р., etal., Эффекты Echinaforce® лечения exvivo-стимулированных клеток крови.
Phytomedicine, 2011.18 (10): p.826-31.
12. Sharma, M., Etal., Индукциямножественныхпровоспалительныхцитокинов респираторнымивирусамии обратное действие
стандартизированнымиEchinacea, a сильнодействующий противовирусный растительный экстракт. Antiviral Res, 2009.83 (2): p.165-70.
13. Sharma, SM, etal., Бактерицидные
(Echinaforce): «Двойные» действия против дыхательных путей. бактерии.Phytomedicine, 2010.17 (8-9): p.563-8.
14. Шарма, М., Р. Шоп, и Дж. Б. Хадсон, «Эхинацея» как противовоспалительный агент: «влияние»
физиологически значимых параметров. PhytotherRes, 2009.23 (6): p.863-7.
15. Pleschka, S., Etal., Антивирусныесвойстваирежимдействиястандартизированного экстракта эхинацеиpurpurea
против высокопатогенныхavian «Вирус гриппа» (H5N1, «H7N7)» и «винное происхождение» h2N1 (S-OIV). «Вирол» J,
2009. №6: стр.197.
16. Шарма, M., R.Schoop, иJBHudson, ЭффективностьEchinaceaina3-Dtissuemodel ofhuman
airwayepithelium.PhytotherRes, 2010.24 (6): p.900-4.
17. Canlas, J., Etal., Echinaceaandtrypanasomatidparasite взаимодействия: ингибиторы роста иanti-
воспалительные эффекты Echinacea.Pharm Биол, 2010. №48 (9): p.1047-52.
18. Woelkart, K., Etal., Биодоступностьифармакокинетикапрепаратов эхинацеиpurpurea и их
взаимодействия с иммунной системой.IntJClinPharmacolTher, 2006.44 (9): p.401-8.
19. Sharma, M., Etal., Возможное использование эхинацеи при акне: контроль за ростом Propionibacteriumacnes
и воспалением. Res, 2011.25 (4): p.517-21.
20. Gertsch, J., Etal., Эхинацеиалкиламиды модулируютTNF-альфа ген экспрессию через каннабиноидный рецептор
CB2 и несколько путей передачи сигналов .FEBSLett, 2004.577 (3): p.563-9.
21. Vimalanathan, S., Etal., Предупреждениевирус гриппаиндуцированныйбактериальнойсуперинфекцией стандартизованным
Echinaceapurpurea, посредством регуляции поверхности экспрессия рецепторов в человеческих, бронхиальных, эпителиальных клетках.
VirusRes, 2017.233: p.51-59.
22. Вималанатан, S., R.Schoop, и J.Hudson, Высокоэффективная противогриппозная терапия от
Echinacea purpureafresh «Травы» и «корни» настойки.
1-5.
23. Brinkeborn, RM, DVShah, иFHDegenring, Echinaforce и другое растение Echinaceafresh холодное. рандомизированное, плацебо-контролируемое, двойное слепое
клиническое исследование.Phytomedicine, 1999.6 (1): p.1-6.
24. Jawad, M., Etal., Безопасностьи ЭффективностьПрофильEchinaceapurpureatoPreventCommonColdEpisodes: A
Randomized, Double -Слепой, «Испытание с плацебо-контролем». «Эвид», «Дополнение», «Альтернат», «2012», «2012»:
p. 84 1315.
25. Raduner, S., Etal., Алкиламиды изEchinaceaarea новый класс каннабиномиметиков. Cannabinoidtype
2receptor-зависимыеand -независимые иммуномодулирующие эффекты. JBiolChem, 2006.№281 (20): стр.
14192-206.
Ингибирование IFN-γ-зависимой противовирусной эпителиальной защиты дыхательных путей сигаретным дымом | Респираторные исследования
Гюркан Ф., Кирал А., Дагли Э., Каракоч Ф: Влияние пассивного курения на развитие респираторно-синцитиального вирусного бронхиолита. Eur J Epidemiol 2000, 16: 465–468.
Артикул PubMed Google ученый
Bradley JP, Bacharier LB, Bonfiglio J, Schechtman KB, Strunk R, Storch G, Castro M: Тяжесть респираторно-синцитиального вирусного бронхиолита зависит от воздействия сигаретного дыма и атопии. Педиатрия 2005, 115: e7-e14.
PubMed Google ученый
Министерство здравоохранения и социальных служб США: Последствия для здоровья непроизвольного воздействия табачного дыма: отчет Главного хирурга. Департамент здравоохранения и социальных служб, Центры по контролю и профилактике заболеваний, Координационный центр по укреплению здоровья, Национальный центр профилактики хронических заболеваний и укрепления здоровья, Управление по курению и здоровью, Атланта, Джорджия; 2006.
Benseñor IM, Cook NR, Lee IM, Chown MJ, Hennekens CH, Buring JE, Manson JE: Активное и пассивное курение и риск простуды у женщин. Ann Epidemiol 2001, 11: 225–231.
Артикул PubMed Google ученый
Блейк Г. Х., Абелл Т. Д., Стэнли РГ: Курение сигарет и инфекции верхних дыхательных путей среди новобранцев, проходящих базовую боевую подготовку. Ann Intern Med 1988, 109: 198–202.
CAS Статья PubMed Google ученый
Коэн С., Тиррелл Д.А., Рассел М.А., Джарвис М.Дж., Смит А.П.: Курение, употребление алкоголя и предрасположенность к простуде. Am J Public Health 1993, 83: 1277–1283.
CAS Статья PubMed PubMed Central Google ученый
Карк Д.Д., Лебиуш М., Раннон L: Курение сигарет как фактор риска эпидемического гриппа A (h2N1) у молодых мужчин. N Engl J Med 1982, 307: 1042–1046.
CAS Статья PubMed Google ученый
Бах Э.А., Агует М., Шрайбер RD: Рецептор IFN-g: парадигма передачи сигналов рецептора цитокина. Annu Rev Immunol 1997, 15: 563–591.
CAS Статья PubMed Google ученый
Танигучи Т., Такаока A: Слабый сигнал для сильных ответов: снова интерферон-α / β. Nature Reviews 2001, 2: 378–386.
CAS Статья PubMed Google ученый
Bach EA, Tanner JW, Marsters S, Ashkenazi A, Aguet M, Shaw AS, Schreiber RD: Лиганд-индуцированная сборка и активация рецептора гамма-интерферона в интактных клетках. Mol Cell Biol 1996, 16: 3214–3221.
CAS Статья PubMed PubMed Central Google ученый
Greenlund AC, Morales MO, Viviano BL, Yan H, Krolewski J, Schreiber RD: Рекрутмент статов тирозин-фосфорилированными цитокиновыми рецепторами: упорядоченный обратимый процесс, управляемый аффинностью. Иммунитет 1995, 2: 677–687.
CAS Статья PubMed Google ученый
Look DC, Pelletier MR, Tidwell RM, Roswit WT, Holtzman MJ: Stat1 зависит от транскрипционной синергии с Sp1. J Biol Chem 1995, 270: 30264-30267.
CAS Статья PubMed Google ученый
Zhang JJ, Vinkemeier E, Gu W, Chakravarti D, Horvath CM, Darnell JE: Две контактные области между Stat1 и CBP / p300 в передаче сигналов интерферона g. Proc Natl Acad Sci USA 1996, 93: 15092–15096.
CAS Статья PubMed PubMed Central Google ученый
Joseph TD, Look DC: Специфическое ингибирование путей передачи сигнала интерферона аденовирусной инфекцией. J Biol Chem 2001, 276: 47136–47142.
CAS Статья PubMed Google ученый
Aldallal N, McNaughton EE, Manzel LJ, Richards AM, Zabner J, Ferkol TW, Look DC: Воспалительная реакция в эпителиальных клетках дыхательных путей, выделенных от пациентов с муковисцидозом. Am J Respir Crit Care Med 2002, 166: 1248–1256.
Артикул PubMed Google ученый
Look DC, Keller BT, Rapp SR, Holtzman MJ: Избирательная индукция молекулы межклеточной адгезии-1 интерфероном-g в эпителиальных клетках дыхательных путей человека. Am J Physiol 1992, 263: L79-L87.
CAS PubMed Google ученый
Frick AG, Joseph TD, Pang L, Rabe AM, St. Geme JW III, Look DC: Haemophilus influenzae стимулирует экспрессию ICAM-1 на клетках респираторного эпителия. J Immunol 2000, 164: 4185–4196.
CAS Статья PubMed Google ученый
Chin CL, Manzel LJ, Lehman EE, Humlicek AL, Shi L, Starner TD, Denning GM, Murphy TF, Sethi S, Look DC: Haemophilus influenzae от пациентов с обострением хронической обструктивной болезни легких вызывают усиление воспаления чем колонизаторы. Am J Respir Crit Care Med 2005, 172: 85–91.
Артикул PubMed PubMed Central Google ученый
Ramaswamy M, Shi L, Monick MM, Hunninghake GW, Look DC: Специфическое ингибирование передачи сигнала интерферона типа I респираторно-синцитиальным вирусом. Am J Respir Cell Mol Biol 2004, 30: 893–900.
CAS Статья PubMed Google ученый
Ramaswamy M, Shi L, Varga SM, Barik S, Behlke MA, Look DC: Неструктурный белок 2 респираторно-синцитиального вируса специфически ингибирует передачу сигнала интерферона I типа. Virology 2006, 344: 328–339.
CAS Статья PubMed Google ученый
Humlicek AL, Manzel LJ, Chin CL, Shi L, Excoffon KJ, Winter MC, Shasby DM, Look DC: Параклеточная проницаемость ограничивает эпителиальные реакции дыхательных путей, что позволяет избирательно активировать медиаторы на базолатеральной поверхности. J Immunol 2007, 178: 6395–6403.
CAS Статья PubMed Google ученый
Manzel LJ, Chin CL, Behlke MA, Look DC: Регуляция индуцированной бактериями молекулы-1 межклеточной адгезии с помощью CCAAT / связывающих белков энхансера. Am J Respir Cell Mol Biol 2009, 40: 200–210.
CAS Статья PubMed Google ученый
Расбанд WS: ImageJ. [http://rsb.info.nih.gov/ij/] Национальные институты здравоохранения, Бетесда, Мэриленд; 1997.
Хассард TH: Понимание биостатистики. Сент-Луис, Миссури: Мосби-Год Бук, Инк; 1991.
Google ученый
Veals SA, Schindler C, Leonard DB, Fu XY, Aebersold RH, Darnell JE, Levy DE: Субъединица фактора транскрипции, чувствительного к альфа-интерферону, связана с регуляторным фактором интерферона и семействами ДНК Myb. связывающие белки. Mol Cell Biol 1992, 12: 3315–3324.
CAS Статья PubMed PubMed Central Google ученый
Wong LH, Hatzinisiriou I, Devenish RJ, Ralph SJ: Праймирование IFN-g активирует компоненты фактора 3 гена, стимулированного IFN (ISGF3), повышая чувствительность IFN-устойчивых клеток меланомы к IFN типа I. J Immunol 1998, 160: 5475–5484.
CAS PubMed Google ученый
Look DC, Pelletier MR, Holtzman MJ: Селективное взаимодействие подмножества белков, связывающих элемент ответа на интерферон-g, с промотором гена молекулы межклеточной адгезии-1 (ICAM-1) контролирует характер экспрессии на эпителиальных клетках. J Biol Chem 1994, 269: 8952–8958.
CAS PubMed Google ученый
Horvath CM, Darnell JE: Антивирусное состояние, индуцированное альфа- и гамма-интерфероном, требует транскрипционно активного белка Stat1. J Virol 1996, 70: 647–650.
CAS PubMed PubMed Central Google ученый
Walter MJ, Look DC, Tidwell RM, Roswit WT, Holtzman MJ: Целевое ингибирование интерферон-γ-зависимой экспрессии ICAM-1 с использованием доминантно-отрицательного Stat1. J Biol Chem 1997, 272: 28582–28589.
CAS Статья PubMed Google ученый
Бухвалова М.С., Принц Г.А., Бланко JCG: Респираторно-синцитиальный вирус инфицирует и преждевременно реплицируется в легких, несмотря на ранее существовавший иммунитет. J Virol 2007, 81: 9443–9450.
CAS Статья PubMed PubMed Central Google ученый
Pryor WA, Prier DG, Church DF: Исследование электронно-спинового резонанса основного и побочного сигаретного дыма: природа свободных радикалов в газовой фазе дыма и в сигаретной смоле. Environ Health Perspect 1983, 47: 345–355.
CAS Статья PubMed PubMed Central Google ученый
Прайор В.А., Стоун К: Окислители в сигаретном дыме. Радикалы, перекись водорода, пероксинитрат и пероксинитрит. Ann N Y Acad Sci 1993, 686: 12–28.
CAS Статья PubMed Google ученый
Eiserich JP, Vliet A, Handelman GJ, Halliwell B, Cross CE: Диетические антиоксиданты и биомолекулярное повреждение, вызванное сигаретным дымом: сложное взаимодействие. Am J Clin Nutr 1995, 62: 1490S-1500S.
CAS PubMed Google ученый
Rahman I, Biswas SK, Kode A: Окислительный и антиоксидантный баланс в дыхательных путях и заболеваниях дыхательных путей. Eur J Pharmacol 2006, 533: 222–239.
CAS Статья PubMed Google ученый
Zang LY, Stone K, Pryor WA: Обнаружение свободных радикалов в водных экстрактах сигаретной смолы с помощью электронно-спинового резонанса. Free Radic Biol Med 1995, 19: 161–167.
CAS Статья PubMed Google ученый
MacNee W: Окислители / антиоксиданты и ХОБЛ. Комод 2000, 117: 303S-317S.
CAS Статья PubMed Google ученый
Джоши-Барве С., Аманчерла К., Патил М., Бхатнагар А., Мэтьюз С., Гобеджишвили Л., Кейв М., Макклейн С., Барве С. Акролеин, повсеместно распространенный загрязнитель и продукт гидропероксида липидов, ингибирует антивирусную активность. интерферон-альфа: отношение к гепатиту С. Free Radic Biol Med 2009, 47: 47–54.
CAS Статья PubMed PubMed Central Google ученый
Groskreutz DJ, Monick MM, Babor EC, Nyunoya T, Varga SM, Look DC, Hunninghake GW: Сигаретный дым изменяет индуцированный респираторно-синцитиальным вирусом апоптоз и репликацию. Am J Respir Cell Mol Biol 2009, 41: 189–198.
CAS Статья PubMed PubMed Central Google ученый
Hellermann GR, Nagy SB, Kong X, Lockey RF, Mohapatra SS: Механизм острой воспалительной реакции, вызванной конденсатом сигаретного дыма, в эпителиальных клетках бронхов человека. Respir Res 2002, 3: 22.
Статья PubMed PubMed Central Google ученый
Лю Х, Того С., Аль-Муготир М., Ким Х., Фанг Q, Кобаяши Т., Ван Х, Мао Л., Биттерман П., Реннард С.: NF-kappaB обеспечивает выживание эпителиальных клеток бронхов человека. подвергается воздействию экстракта сигаретного дыма. Respir Res 2008, 9: 66.
Статья PubMed PubMed Central Google ученый
Petecchia L, Sabatini F, Varesio L, Camoirano A, Usai C, Pezzolo A, Rossi GA: Повреждение эпителиальных клеток бронхиальных дыхательных путей после воздействия сигаретного дыма включает разборку компонентов плотного соединения, опосредованную внеклеточным сигналом. регулируемый путь киназы 1/2. Сундук 2009, 135: 1502–1512.
CAS Статья PubMed Google ученый
Shapiro SD: Дым попадает в ваши камеры. Am J Respir Cell Mol Biol 2004, 31: 481–482.
CAS Статья PubMed Google ученый
Моррисон Д., Рахман И., Ланнан С., Макни W: Проницаемость эпителия, воспаление и окислительный стресс в воздушном пространстве курильщиков. Am J Respir Crit Care Med 1999, 159: 473–479.
CAS Статья PubMed Google ученый
Рахман И., Макни W: Регулирование уровней окислительно-восстановительного глутатиона и транскрипции генов при воспалении легких: терапевтические подходы. Free Radic Biol Med 2000, 28: 1405–1420.
CAS Статья PubMed Google ученый
Rahman I, Smith CAD, Lawson MF, Harrison DJ, MacNee W: Индукция γ-глутамилцистеинсинтетазы сигаретным дымом связана с AP-1 в альвеолярных эпителиальных клетках человека. FEBS Lett 1996, 396: 21–25.
CAS Статья PubMed Google ученый
Knörr-Wittmann C, Hengstermann A, Gebel S, Alam J, Müller T: Характеристика активации Nrf2 и экспрессии гемоксигеназы-1 в клетках NIh4T3, подвергнутых воздействию водных экстрактов сигаретного дыма. Free Radic Biol Med 2005, 39: 1438–1448.
Артикул PubMed Google ученый
HuangFu WC, Liu J, Harty RN, Fuchs SY: Продукты для курения сигарет подавляют противовирусные эффекты интерферона типа I посредством зависимой от фосфорилирования регуляции его рецептора. FEBS Lett 2008, 582: 3206–3210.
CAS Статья PubMed PubMed Central Google ученый
Goh KC, Haque SJ, Williams BRG: Киназа p38 MAP необходима для фосфорилирования серина STAT1 и активации транскрипции, индуцированной интерферонами. EMBO J 1999, 18: 5601–5608.
CAS Статья PubMed PubMed Central Google ученый
Нгуен Х., Рамана С.В., Байес Дж., Старк GR: Роль фосфатидилинозитол-3-киназы в интерферон-γ-зависимом фосфорилировании STAT1 по серину 727 и активации экспрессии генов. J Biol Chem 2001, 276: 33361–33368.
CAS Статья PubMed Google ученый
Deb DK, Sassano A, Lekmine F, Majchrzak B, Verma A, Kambhampati S, Uddin S, Rahman A, Fish EN, Platanias LC: Активация протеинкиназы Cδ с помощью IFN-γ. J Immunol 2003, 171: 267–273.
CAS Статья PubMed Google ученый
Chang Y, Holtzman MJ, Chen C: Дифференциальная роль киназ семейства janus (JAK) в индуцированной интерфероном-γ экспрессии легочного эпителия ICAM-1: включая белковые взаимодействия между JAK, фосфолипазой cγ, c-Src и STAT1. Mol Pharmacol 2004, 65: 589–598.
CAS Статья PubMed Google ученый
Bauer CM, Dewitte-Orr SJ, Hornby KR, Zavitz CC, Lichty BD, Stämpfli MR, Mossman KL: Сигаретный дым подавляет опосредованный интерфероном антивирусный иммунитет в фибробластах легких и эпителиальных клетках легких. J Interferon Cytokine Res 2008, 28: 167–179.
CAS Статья PubMed Google ученый
Mian MF, Stämpfli MR, Mossman KL, Ashkar AA: Ослабление в сигаретном дыме индуцированных поли I: C врожденных противовирусных реакций в PBMC человека в основном связано с ингибированием продукции IFN-бета. Mol Immunol 2009, 46: 821–829.
CAS Статья PubMed Google ученый
Браун К.М., Корниш Т., Валм А., Кундифф Дж., Поли Дж. Л., Фан С: Иммунотоксикология конденсатов сигаретного дыма: подавление реакции макрофагов на гамма-интерферон. Toxicol Appl Pharmacol 1998, 149: 136–143.
CAS Статья PubMed Google ученый
Эдвардс К., Браун К.М., Эванс Г., Сурека А.О., Вентилятор S: Конденсаты сигаретного дыма в основном и побочном потоке подавляют реакцию макрофагов на гамма-интерферон. Hum Exp Toxicol 1999, 18: 233–240.
CAS Статья PubMed Google ученый
Диллон Н.К., Мерфи В.Дж., Филла МБ, Креспо А.Дж., Латам Х.А., О’Брайен-Ладнер А: Понижающая модуляция передачи сигналов IFN-гамма в альвеолярных макрофагах, выделенных от курильщиков. Toxicol Appl Pharmacol 2009, 237: 22–28.
CAS Статья PubMed PubMed Central Google ученый
Ван Г., Забнер Дж., Диринг С., Лаунспач Дж., Шао Дж., Боднер М., Джолли Д. Д., Дэвидсон Б. Л., МакКрей PBJ: Повышение проницаемости эпителиальных соединений увеличивает перенос генов в эпителий дыхательных путей In vivo. Am J Respir Cell Mol Biol 2000, 22: 129–138.
Артикул PubMed Google ученый
Дранник А.Г., Пулади М.А., Роббинс С.С., Гончарова С.И., Кианпур С., Штемпфли МР: Влияние сигаретного дыма на клиренс и воспаление после инфекции Pseudomonas aeruginosa. Am J Respir Crit Care Med 2004, 170: 1164–1171.
Артикул PubMed Google ученый
Sopori M: Влияние сигаретного дыма на иммунную систему. Nat Rev Immunol 2002, 2: 372–377.
CAS Статья PubMed Google ученый
Роббинс С.С., Бауэр С.М., Вуйчич Н., Гашлер Г.Дж., Лихти Б.Д., Браун Е.Г., Штемпфли MR: Сигаретный дым влияет на иммунные воспалительные реакции на грипп у мышей. Am J Respir Crit Care Med 2006, 174: 1342–1351.
CAS Статья PubMed Google ученый
Vlahos R, Bozinovski S, Jones JE, Powell J, Gras J, Lilja A, Hansen MJ, Gualano RC, Irving L, Anderson GP: Профили дифференцированной протеазы, врожденного иммунитета и индукции NF-κB во время воспаление легких, вызванное субхроническим воздействием сигаретного дыма у мышей. Am J Physiol Lung Cell Mol Physiol 2006, 290: L931-L945.
CAS Статья PubMed Google ученый
Birrell MA, Wong S, Catley MC, Belvisi MG: Влияние табачного дыма на ключевые сигнальные пути врожденного иммунного ответа в макрофагах легких. J Cell Physiol 2007, 214: 27–37.
Артикул Google ученый
Devalia JL, Campbell AM, Sapsford RJ, Rusznak C, Quint D, Godard P, Bousquet J, Davies RJ: Влияние диоксида азота на синтез воспалительных цитокинов, экспрессируемых эпителиальными клетками бронхов человека in vitro. Am J Respir Cell Mol Biol 1993, 9: 271–278.
CAS Статья PubMed Google ученый
Rusznak C, Devalia JL, Sapsford RJ, Davies RJ: Озон-индуцированное высвобождение медиатора из эпителиальных клеток бронхов человека in vitro и влияние недокромила натрия. Eur Respir J 1996, 9: 2298–2305.
CAS Статья PubMed Google ученый
Роббинс С.С., Доу Д.Е., Гончарова С.И., Пулади М.А., Дранник А.Г., Свирски Ф.К., Кокс Г., Штемпфли MR: Сигаретный дым уменьшает количество дендритных клеток легких и влияет на противовирусный иммунный ответ. Am J Respir Cell Mol Biol 2004, 30: 202–211.
CAS Статья PubMed Google ученый
Castro SM, Kolli D, Guerrero-Plata A, Garofalo RP, Casola A: Конденсат сигаретного дыма усиливает индуцированное респираторно-синцитиальным вирусом высвобождение хемокинов, модулируя NF-каппа B и активацию регуляторного фактора интерферона. Toxicol Sci 2008, 106: 509–518.
CAS Статья PubMed PubMed Central Google ученый
Phaybouth V, Wang SZ, Hutt JA, McDonald JD, Harrod KS, Barrett EG: Сигаретный дым подавляет выработку цитокинов Th2 и увеличивает экспрессию RSV в модели новорожденных. Am J Physiol Lung Cell Mol Physiol 2006, 290: L222-L231.
CAS Статья PubMed Google ученый
% PDF-1.7 % 178 0 объект > эндобдж xref 178 100 0000000016 00000 н. 0000003092 00000 н. 0000003302 00000 н. 0000003431 00000 н. 0000003467 00000 н. 0000004019 00000 п. 0000004094 00000 н. 0000004232 00000 н. 0000004369 00000 п. 0000004507 00000 н. 0000004644 00000 п. 0000004782 00000 н. 0000004919 00000 н. 0000005058 00000 н. 0000005201 00000 н. 0000005485 00000 н. 0000005599 00000 н. 0000005711 00000 н. 0000005988 00000 н. 0000006372 00000 н. 0000006663 00000 н. 0000007096 00000 н. 0000007515 00000 н. 0000007552 00000 н. 0000008399 00000 н. 0000008542 00000 н. 0000008675 00000 н. 0000008807 00000 н. 0000009056 00000 н. 0000009486 00000 н. 0000009936 00000 н. 0000010533 00000 п. 0000011137 00000 п. 0000011415 00000 п. 0000011442 00000 п. 0000011753 00000 п. 0000012031 00000 н. 0000012058 00000 п. 0000012751 00000 п. 0000013730 00000 п. 0000014597 00000 п. 0000015348 00000 п. 0000016291 00000 п. 0000016460 00000 п. 0000016906 00000 п. 0000017963 00000 п. 0000018106 00000 п. 0000019150 00000 п. 0000019780 00000 п. 0000022430 00000 н. 0000022500 00000 п. 0000030441 00000 п. 0000037775 00000 п. 0000074987 00000 п. 0000080868 00000 п. 0000080957 00000 п. 0000081038 00000 п. 0000085358 00000 п. 0000085621 00000 п. 0000085797 00000 п. 0000121258 00000 н. 0000121846 00000 н. 0000127943 00000 н. 0000128205 00000 н. 0000128320 00000 н. 0000128420 00000 н. 0000128505 00000 н. 0000136611 00000 н. 0000136894 00000 н. 0000137288 00000 н. 0000137315 00000 н. 0000137829 00000 н. 0000146133 00000 п. 0000146411 00000 н. 0000146822 00000 н. 0000146849 00000 н. 0000147378 00000 н. 0000147466 00000 н. 0000154029 00000 н. 0000154312 00000 н. 0000154646 00000 н. 0000154673 00000 н. 0000155123 00000 н. 0000158410 00000 н. 0000158691 00000 н. 0000166269 00000 н. 0000166551 00000 н. 0000169157 00000 н. 0000169437 00000 н. 0000178580 00000 н. 0000178863 00000 н. 0000179134 00000 н. 0000179445 00000 н. 0000179595 00000 н. 0000179728 00000 н. 0000179878 00000 н. 0000180029 00000 н. 0000180180 00000 н. 0000180330 00000 н. 0000002296 00000 н. трейлер ] / Назад 619219 >> startxref 0 %% EOF 277 0 объект > поток hb«b`
Экстракт Rhododendron album Blume ингибирует TNF-α / IFN-γ индуцированную продукцию хемокинов посредством блокады NF-κB и активации JAK / STAT в эпидермальных кератиноцитах человека
Введение
Атопический дерматит (АД) — хронический воспалительный кожное заболевание и стойкое воспалительное заболевание кожи, сопровождающееся экзематозными поражениями и сильным зудом.Это вызвано экологические и генетические факторы, включая микробную инфекцию и загрязнители окружающей среды (1-3). БА возникает в результате сочетания сильного зуда и эпидермального барьера. аномалии, несбалансированные иммунные ответы и генетические предрасположенность (4,5). Диагноз БА основывается на следующие клинические фенотипы: эритематозные папулы, экзематозные кожа и сильный зуд (6,7).
Хемокины — это большая группа малых цитокинов. продуцируются различными типами клеток и разделены на CX3C, Подсемейства CXC, CC и C на основе Nh3-терминала цистеин-мотивы.На основе категоризации этих подсемейства, методическая номенклатурная система хемокинов лиганды были разработаны в предыдущих исследованиях (8). Основная функция хемокинов: контроль рекрутирования воспалительных клеток, в том числе макрофаги, Т-клетки, эозинофилы и трафик дендритных клетки (ДК) в очагах воспаления и инфекции (9). Тимус и активация регулируются хемокины (TARC / CCL17) — это хемокины CC, которые конститутивно экспрессируется и продуцируется кератиноцитами, происходящими из моноцитов, и дендритные клетки (10,11).Хемокины, полученные из макрофагов (MDC / CCL22) также являются специфическими лигандами для хемокинов C-C. рецептор типа 4 (CCR4). MDC / CCL22 постоянно производится эпителиальные клетки, В-клетки, кератиноциты, макрофаги и дендритные клетки (10,12). MDC и TARC могут служить важным функция в увеличении заболеваемости некоторыми кожными заболеваниями, в том числе AD. Предыдущие исследователи сообщали, что сыворотка концентрации MDC и TARC положительно коррелируют с тяжесть заболевания у пациентов с БА (13).Учитывая все эти факторы, MDC и TARC может участвовать в патогенезе AD.
Фактор некроза опухоли (TNF) является провоспалительным цитокин с множеством биологических функций, включая цитостатическое и цитотоксические эффекты, дифференциация и пролиферация в различных типы опухолевой клетки. Противоопухолевые эффекты TNF усиливаются за счет интерфероны (ИФН) (14). В фактор ядерной транскрипции, ядерный фактор (NF) -κB был сообщается, что это антиапоптотический фактор, который служит основным функция выживания клеток во время апоптоза, индуцированного цитокинами, включая TNF-α / IFN-γ и различные химиотерапевтические агенты (15,16).Кроме того, фосфорилирование киназа Janus / сигнальные преобразователи и активаторы транскрипции (JAK / STAT) путь, как сообщается, является важным противовоспалительный фактор. Активация JAK стимулирует клетку дифференцировка, пролиферация, миграция клеток и апоптоз. Кимура и др. (14) сообщили, что комбинация лечения TNF-α и IFN-γ привела к в усиленном иммунном ответе фибробластов мышей (14). Кроме того, этот TNF-α / IFN-γ комбинированное лечение также увеличивало in vitro и in vivo индукция воспаления у человека (17,18).На основании этих наблюдений наши группа предположила, что синергизм атопического дерматита и иммунные ответы приводят к подавлению TNF-α / IFN-γ-индуцированные механизмы выживания клеток.
Альбом рододендронов Blume (RA) — вид Растение, принадлежащее к семейству вересковых. В первую очередь встречается во влажных первобытных лесах на больших высотах по всей западной и Центральная Ява. РА 1-2 м высотой, а листья (5-12 см длиной и 2-3 см шириной) жесткие, с коричневой чешуйчатой нижней стороной.Венчик широко колокольчатая, бледно-желтого цвета с разбросанными коричневые чешуйки. РА является эндемичным для Явы и редко встречается в дикой природе. Однако РА широко выращивают в питомниках по всему миру. особенно в США. Некоторые растения, относящиеся к рода Rhododendron документально подтвердили противораковые (19,20) и антиоксидантные функции (21). Следовательно, РА может также оказывать антиатопическая или противовоспалительная активность. Настоящее исследование сосредоточены на противовоспалительном действии метанола RA (MeOH) извлечь (РАМЭ) (22).В настоящий момент, Нет данных о противовоспалительной активности этого растения. Таким образом, целью настоящего исследования было оценить противовоспалительная и антиатопическая дерматитная активность RAME в Клетки кератиноцитов HaCaT.
Материалы и методы
Приготовление экстракта RA
Вид растений был собран из Кантиги в Индонезии в 2008 году, и он был определен Центром Фармацевтические и медицинские технологии, Агентство по оценке и Применение технологии (Тангеранг, Индонезия) и подтверждено Гербарий Bogoriense (Индонезийский институт наук, Богор, Индонезия).Образец ваучера (KRIB 0019989) депонирован в гербарий Корейского научно-исследовательского института биологических наук и Биотехнологии, а также в Центре фармацевтики и Медицинские технологии и гербарий Bogoriense. Растительный материал РА обрабатывали МеОН и обрабатывали ультразвуком в течение 15 мин при 36,5 кГц, используя Ванна для ультразвуковой очистки (Branson Ultrasonics; Emerson Electric Co., Сент-Луис, Миссури, США) и инкубировали в течение 2 ч при комнатной температуре. Эту процедуру повторяли 30 раз в течение 3 дней для получения извлекать.Метанольный экстракт (360 мг) суспендировали в дистиллированной воды, добавляли и перемешивали такое же количество н-гексана, а затем растворимая в н-гексане фракция и водорастворимая фракция были разделены. Это было выполнено трижды, затем вата. фильтрация и концентрирование при пониженном давлении для получения Фракция н-гексана (выход 17,2 мг). Затем фракция н-гексана была удалили, и такое же количество хлороформа было добавлено к оставшийся водный слой. Фракция хлороформа (выход 21.0 мг) было получен таким же образом. Такое же количество этилацетата было позже добавляется к оставшейся воде, и фракция этилацетата (выход 14,0 мг) получали таким же образом. Наконец, в водный слой добавляли эквивалентное количество бутанола, и бутанольную фракцию (выход 47,3 мг) получали таким же способом. Оставшийся водный слой затем концентрировали для получения водного фракция (выход 235,5 мг).
Культура клеток
Клеточная линия кератиноцитов НаСаТ человека (АТСС; Американская коллекция типовых культур, Манассас, Вирджиния, США) были выращены в среде Игла, модифицированной Дульбекко с высоким содержанием глюкозы (DMEM) с добавлением 10% фетальной телячьей сыворотки (FBS) и антибиотиков (100 Ед / мл пенициллина и 100 мкг / мл стрептомицина) при 37 ° C в увлажненный инкубатор с 5% CO2.До дальнейшего в ассоциативных исследованиях питательную среду заменили на бессывороточную Средняя. Клетки контрольной группы обрабатывали диметил только сульфоксид (ДМСО, 0,1% на 100 мкл). DMEM, FBS, пенициллин, стрептомицин и PBS (pH 7,4) были приобретены в Гибко; Thermo Fisher Scientific, Inc. (Уолтем, Массачусетс, США). Рекомбинантный TNF-α (10 нг / мл) был приобретен у Invitrogen; Термо Fisher Scientific, Inc. и IFN-γ (10 нг / мл) был приобретен у Merck KGaA (Дармштадт, Германия). Клетки HaCaT были предварительно обработаны. с РАМЭ (2.5, 5, 10, 20 мкг / мл) в течение 1 ч, а затем обрабатывали TNF-α (10 нг / мл) и IFN-γ (10 нг / мл) и инкубировали в течение 24 часов при 37 ° C. В другом наборе культур клетки были совместно инкубировали с 5 мкМ Bay11-7082, ядерный фактор κ усилитель гена легкого полипептида в ингибиторе B-клеток, α (IκB-α) ингибитор, 10 мкМ SB203580, ингибитор p38 и 10 мкМ SP600125, ингибитор N-концевой киназы c-Jun (JNK) (Calbiochem; EMD Millipore, Биллерика, Массачусетс, США) в течение 1 ч при 37 ° C в увлажненном помещении. Инкубатор с 5% CO2.
Жизнеспособность клеток
Чтобы подтвердить действие RAME на клетки HaCaT, снижение MTT (Amresco, Inc., Солон, Огайо, США) жизнеспособными клетками был измерен. Клетки высевали в 96-луночные культуральные планшеты (SPL Науки о жизни, Почхон, Корея) с плотностью 1 × 104 клеток / лунку, и давали возможность связываться в течение 6 часов. Клеткам разрешили прикрепить в течение 6 часов, и культивирование продолжали в течение 24 часов после добавление РАМЭ (2,5, 5, 10, 20 мкг / мл) при 37 ° C. Клетки культивировали с раствором МТТ 0,5 мг / мл. Через 4 часа инкубации при 37 ° C в 5% CO2 супернатант удаляли и ДМСО был добавлен. Поглощение измеряли при 570 нм с помощью микропланшета. ридер (Tecan Group, Ltd., Меннедорф, Швейцария). Процент жизнеспособных клеток рассчитывали, беря оптическую плотность клетки после определенного лечения и делящие это число с оптической плотностью необработанных контрольных клеток с последующим умножение на 100.
Интерлейкин (ИЛ) -6 и ИЛ-8 производство
Питательная среда была собрана, и производство цитокинов (IL-6 и IL-8) в супернатанте измеряли с использованием Наборы для ELISA (IL-6; номер по каталогу 555220, BD Biosciences, Сан-Хосе, Калифорния, СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ; Ил-8; Кот.нет. DY208, R&D Systems, Inc., Миннеаполис, Миннесота, США) согласно протоколам производителей, как и ранее описано (23,24).
Производство TARC и MDC
Культуральную среду клеток собирали и продукцию хемокинов (TARC и MDC) в супернатанте измеряли. с использованием наборов ELISA (каталожные номера TARC, каталожный номер DY364 и MDC, каталожный номер. DY336; R&D Systems, Inc.) по данным производителя. протокол.
Обратная транскрипция-количественная анализ полимеразной цепной реакции (RT-qPCR)
Тотальную РНК экстрагировали с использованием реагента TRIzol (Thermo Fisher Scientific, Inc.) для анализа RT-qPCR. кДНК была синтезирована с использованием набора для синтеза кДНК AMPIGENE® (кат. ENZ-KIT106; Enzo Life Sciences, Inc., Фармингдейл, Нью-Йорк, США). КПЦР выполняли с использованием SYBR Green PCR Master Mix (KAPA Наборы SYBR® FAST qPCR; Kapa Biosystems, Inc., Уилмингтон, Массачусетс, США). Реакции проводили в системе ПЦР в реальном времени CFX96. (Bio-Rad Laboratories, Inc., Геркулес, Калифорния, США), используя следующие условия в термоциклере: 1 этап, 50 ° C в течение 2 минут и 95 ° C в течение 10 минут. мин; стадия 2, 95 ° C в течение 15 секунд и 60 ° C в течение 1 мин.2 этап был повторяется 40 циклов. Относительные уровни мРНК рассчитывали с использованием метод сравнительного порогового цикла 2 — ΔΔCq и нормализовано к GAPDH (25). Последовательности праймеров, используемые в Настоящие исследования перечислены в Таблице I.
Таблица IПраймерные последовательности. |
Таблица I
Праймерные последовательности.
| Ген | Праймер последовательностиa (5′-3 ‘) | Размер фрагмента (бп) |
|---|---|---|
| TARC | 222 | |
| Вперед | CACGCAGCTCGAGGGACCAATGTG | |
| Задний ход | TCAAGACCTCTCAAGGCTTTGCAGG | |
| MDC | 362 | |
| Вперед | AGGACAGAGCATGGCTCGCCTACAGA | |
| Задний ход | TAATGGCAGGGAGGTAGGGCTCCTGA | |
| IL-6 | 124 | |
| Вперед | GACAGCCACTCACCTCTTCA | |
| Задний ход | AGTGCCTCTTTGCTGCTTTC | |
| IL-8 | 299 | |
| Вперед | ATGACTTCCAAGCTGGCCGTGGCT | |
| Задний ход | TTATGAATTCTCAGCCCTCTTCAAAAA | |
| β-актин | 250 | |
| Вперед | CATGTACGTTGCTATCCAGGC | |
| Задний ход | CTCCTTAATGTCACGCACGAT |
Натрия додецилсульфат-полиакриламидный гель электрофорез (SDS-PAGE) и вестерн-блоттинг
Вкратце, клетки HaCaT промывали холодным PBS. а затем лизировали в буфере RIPA (ELPIS Biotech, Inc., Тэджон, Корея) на льду 20 мин при сильном вихре. Лизат клеток был центрифугировали при 10000 × g в течение 10 мин при 4 ° C и белке концентрации были измерены с использованием метода бицинхониновой кислоты. Образцы белков (20 мкг) анализировали на 10% SDS-PADE и электрофоретически перенесенный на поливинилидендифторид (PVDF) мембрана, после чего мембрана PVDF была заблокирована в 5% обезжиренное сухое молоко в трис-буферном физиологическом растворе (TBS, 20 мМ Трис, 0,2 M NaCl, pH 7,5), содержащий 0,05% Tween-20 (TBS / T), в течение 1 ч при комнатной температуре. температура.Мембрану из ПВДФ инкубировали с первичной антитела на ночь при 4 ° C. После трехкратной стирки TBS / T, мембраны инкубировали 1 ч при комнатной температуре. с вторичными антителами, конъюгированными с пероксидазой хрена (HRP), разбавить 1: 5000 5% обезжиренным молоком в TBS / T. Затем мембраны были визуализируется с использованием хемилюминесцентного субстрата SuperSignal West Pico (каталожный номер 32106; Pierce; Thermo Fisher Scientific, Inc.). В Использовались следующие первичные антитела: анти-NF-κB p65 (кат.нет. sc-8242; 1: 1000), анти-JNK (номер по каталогу sc-474; 1: 1000), киназа, регулируемая анти-внеклеточным сигналом (ERK; каталожный номер sc-154; 1: 1000), анти-p38 (каталожный номер sc-7149; 1: 1000), анти-p-p38 (кат. нет. sc-7973; 1: 1000), антифосфорилированный (p-) JAK1 (кат. sc-16773; 1: 1000), анти-JAK1 (номер по каталогу sc-376996; 1: 1000), анти-p-STAT1 (каталожный номер sc-8394; 1: 1000), анти-STAT1 (каталожный номер. sc-464; 1: 1000; все из Santa Cruz Biotechnology, Inc., Даллас, Техас, США), анти-p-ERK 1/2 (каталожный номер 9101; 1: 1000), анти-p-IκB-α (Кот.нет. 2859; 1: 1000), анти-IκB-α (каталожный номер 9242; 1: 1000), анти-NF-κB p-p65 (номер по каталогу 3033; 1: 1000; все из Cell Signaling Technology, Inc., Данверс, Массачусетс, США), анти-p-JNK (каталожный номер. KAP-SA011; 1: 1000; Enzo Life Sciences, Inc.) и анти-β-актин (№ по каталогу 4967; 1: 1000; Cell Signaling Technology, Inc.). В вторичные использованные антитела были следующими: козьи, конъюгированные с HRP анти-кроличий IgG (каталожный номер sc-2030; 1: 5,000 в 5% обезжиренном молоке; Santa Круз Биотехнология, Инк .; используется для обнаружения NF-κB p65, JNK, ERK, p38, p-p38, p-JNK, p-ERK, p-IκB-α, IκB-α, NF-κB, p-p65, β-актин) и конъюгированные с HRP козьи антимышиные IgG (кат.нет. sc-2005; 1: 5,000 в 5% обезжиренном молоке; Санта-Крус биотехнология, инк .; используется для обнаружение JAK1, STAT1, p-JAK1, p-STAT1). Каждый белок был детектируется с помощью системы обнаружения Enhanced Chemiluminescence с использованием ImageQuant LAS4000 (GE Healthcare Life Sciences, Литтл Чалфонт, СОЕДИНЕННОЕ КОРОЛЕВСТВО). Полосы вестерн-блоттинга были количественно определены с использованием программного обеспечения ImageJ. (версия 1.6.0; Национальные институты здравоохранения, Бетесда, Мэриленд, СОЕДИНЕННЫЕ ШТАТЫ АМЕРИКИ).
Анализ люциферазы
клеток HaCaT трансфицировали 0,1 мкг pGL4.32 (luc2P / NF-κB-RE / Hygro, Promega Corporation, Мэдисон, Висконсин, USA) плазмиды с использованием реагента для трансфекции Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific, Inc.) согласно протокол производителя. Затем через 20 ч после трансфекции клетки стимулировали TNF-α / IFN-γ (10 нг / мл) в течение 24 часов при 37 ° C, собирали, а затем оценивали активность люциферазы с помощью Система анализа репортерной люциферазы ONE-Glo ™ (Promega Corporation) согласно протоколу производителя. Нормализация была выполняется путем сравнения с люциферазой Renilla деятельность.
Иммуноцитохимия (ICC)
клеток HaCaT культивировали на предметных стеклах камеры Lab-Tek ™. (Thermo Fisher Scientific, Inc.) и иммобилизовали в этаноле при 4 ° C. на 15 мин. Предметные стекла трижды промывали PBS и блокировали. с 2% (мас. / об.) бычьим сывороточным альбумином (BSA100, Bovogen Biologicals Pty Ltd., Мельбурн, Австралия) в PBS еще на 1 час в комнате температура. Затем слайды инкубировали с субъединицей p65 анти-NF-κB. (каталожный номер sc-8242; кроличьи поликлональные IgG; 1: 200, Санта-Крус. Биотехнология) и анти-STAT1 (каталожный номер sc-464; мышиный моноклональный IgG; 1: 100, Santa Cruz Biotechnology, Inc.) антитела в течение 24 ч при 4 ° С. После трехкратной промывки PBS для удаления излишков первичное антитело, слайды дополнительно инкубировали с Alexa Козьи антикроличьи IgG, конъюгированные с Fluor 488 (кат.нет. А-11034; 1: 1000; Invitrogen; Thermo Fisher Scientific, Inc.) или Texas Red конъюгированные вторичные антитела козы против IgG мыши (кат. А-11003; 1: 1000; Invitrogen; Thermo Fisher Scientific, Inc.) для 2 ч при комнатной температуре, промывают PBS и окрашивают золотом. Реагент антифад, содержащий DAPI (Invitrogen; Thermo Fisher Scientific, Inc.) за 5 минут до позиции и количественная оценка ядер ProLong. Впоследствии слайды были покровные стекла и визуализация с помощью конфокального лазерного сканирования микроскопия (LSM 510 м; Carl Zeiss AG, Оберкохен, Германия).Все образцы были количественно определены по полученным изображениям, которые были взяты при одинаковых условиях воздействия.
Статистический анализ
Данные представляют собой среднее значение ± стандартная ошибка иметь в виду. Статистические различия между группами определялись односторонний дисперсионный анализ с повторными измерениями, за которыми следует Тестирование Стьюдента-Ньюмана-Кеулса в программе SPSS 14.0 (SPSS, Inc., Чикаго, Иллинойс, США). P <0,05 считалось показателем статистически значимая разница.
Результаты
Жизнеспособность клеток
Подтверждено влияние RAME на жизнеспособность клеток. с использованием клеток HaCaT и анализа МТТ.Как показано на рис. 1, RAME не приводил к значительный цитотоксический эффект, не влияющий даже на жизнеспособность клеток при относительно высокой концентрации 20 мкг / мл для период лечения 24 ч. Следовательно, RAME экспериментально был подтверждено, что не проявляет токсичности даже после обработки клеток HaCaT с 20 мкг / мл. Кроме того, действие РАМЭ и растворителя фракция (н-гексан, хлороформ, этилацетат, бутанол, вода слой) на жизнеспособность клеток подтверждали с использованием клеток HaCaT и МТТ анализы. РАМЭ и фракция растворителя не влияли на жизнеспособность клеток, даже при относительно высокой концентрации 20 мкг / мл для 24 ч период лечения (данные не показаны).
Лечение ингибиторами РАМЭ TNF-α / IFN-γ-индуцированная экспрессия TARC и MDC в клетках HaCaT
Далее, эффект подавления TARC и MDC продукция RAME в клетках HaCaT, стимулированных TNF-α / IFN-γ, была исследовали с помощью ELISA и RT-qPCR (рис. 2). TARC и MDC были значительно увеличилось в группе, получавшей TNF-α / IFN-γ по сравнению с необработанная группа. Кроме того, когда клетки HaCaT обрабатывали TNF-α / IFN-γ после обработки RAME, экспрессия мРНК TARC и MDC снижается в зависимости от концентрации, по сравнению с группа, получавшая TNF-α / IFN-γ.Кроме того, лечение RAME подавляет экспрессию белка TARC и MDC в клетках HaCaT стимулировали TNF-α / IFN-γ.
Лечение ингибиторами РАМЭ TNF-α / IFN-γ-индуцированная экспрессия IL-8 и IL-6 в клетках HaCaT
Далее, эффект ингибирования производства ИЛ-6 и ИЛ-8 через лечение RAME были исследованы в HaCaT. клетки стимулировали TNF-α / IFN-γ с помощью ELISA и RT-qPCR (фиг. 3). Результаты показали, что ИЛ-6 и IL-8 были значительно увеличены в группе, получавшей TNF-α / IFN-γ по сравнению с необработанной группой.Кроме того, когда Клетки HaCaT обрабатывали TNF-α / IFN-γ после обработки RAME, Экспрессия мРНК IL-6 и IL-8 снижалась в зависимым от концентрации образом по сравнению с группой, получавшей TNF-α / IFN-γ. Кроме того, лечение RAME подавляло выражение белка IL-6 и IL-8 в клетках HaCaT, стимулированных TNF-α / IFN-γ.
Лечение РАМЭ подавляет активация митоген-активированных протеинкиназ (MAPK) в HaCaT ячейки
TNF-α / IFN-γ активирует экспрессию JNK, ERK и p38 (24).Следовательно, эффект RAME на экспрессию белков JNK, ERK и p38 в клетках HaCaT был исследованы (рис. 4). HaCaT клетки обрабатывали TNF-α / IFN-γ и через 2 часа добавляли RAME. Фосфорилирование изменяется на JNK, ERK и p38, которые являются MAPK, которые известны как важные в пути атопии, были подтверждены. В результаты выявили увеличение фосфорилирования JNK, ERK, и p38 в клетках HaCaT, стимулированных в течение 1 ч TNF-α / IFN-γ, по сравнению с нестимулированными клетками HaCaT. Кроме того, это эксперимент подтвердил, что когда клетки были предварительно обработаны RAME, фосфорилирование JNK, ERK и p38 заметно подавлялось по сравнению с группой, получавшей только TNF-α / IFN-γ.Следовательно, RAME оказывает ингибирующее действие на экспрессию p-JNK, p-ERK и p-p38 после стимуляции TNF-α / IFN-γ.
Рисунок 4Влияние лечения RAME на TNF-α / IFN-γ индуцировали MAPK в клетках HaCaT. Клетки были предварительно обработаны с 2,5, 5,0, 10,0 и 20,0 мкг / мл РАМЭ в течение 1 ч, а затем подвергается воздействию TNF-α и IFN-γ (каждый 10 нг / мл) в течение 2 часов. Экстракты клеток были подготовлены, и активация MAPK была проанализирована с помощью вестерн-блоттинга, с использованием специфических антител.## P <0,01 против отрицательного контроль; * P <0,05, ** P <0,01 и *** P <0,001 по сравнению с клетками, стимулированными TNF-α / IFN-γ. РАМЭ, Экстракт метанола Rhododendron album Blume; TNF-α, опухоль фактор некроза-α; IFN-γ, интерферон-γ; MAPK, митоген-активированный протеинкиназа; п-, фосфорилированный; ERK, внеклеточный сигнально-регулируемая киназа; JNK, N-концевая киназа c-Jun. |
RAME ингибировал активацию NF-κB в Клетки HaCaT
Передача сигналов NF-κB имеет решающее значение при кожном воспалительном AD ответы, индуцированные MDC и TARC (26).Проверить, действительно ли РАМЭ подавление TNF-α / IFN-γ-опосредованной активации NF-κB в кератиноцитов, экспрессия ассоциированных белков была исследовали методом вестерн-блоттинга с использованием специфических антител, ICC, и люциферазные анализы. TNF-α / IFN-γ значительно увеличивал фосфорилирование NF-κB p65 через 1 час. РАМЭ уменьшил фосфорилирование NF-κB p65 в TNF-α / IFN-γ-индуцированных клетках HaCaT (Рис. 5A). TNF-α / IFN-γ-индуцированный ядерная транслокация p65 ингибировалась обработкой RAME при 10 мкг / мл (рис.5Б). Кроме того, ингибирующее действие РАМЭ на TNF-α / IFN-γ-стимулированная ядерная транслокация NF-κB p65 также была наблюдаемые при анализе ICC (рис. 5С). Эти результаты показали, что RAME ингибирует активацию NF-κB в клетках HaCaT.
RAME и Bay11-7082 подавляли активация NF-κB в клетках HaCaT
Субъединица p65 является важным элементом активированного NF-κB. Активация NF-κB в клетках HaCaT, индуцированных TNF-α / IFN-γ, была изучено путем измерения фосфорилирования субъединицы p65 NF-κB с помощью вестерн-блоттинг.Предварительная обработка с помощью RAME и Bay11-7082 снизил уровни p-IκB-α и p-p65 по сравнению с Только лечение TNF-α / IFN-γ, в то время как группа лечения TNF-α / IFN-γ увеличил уровни p-IκB-α и p-p65 по сравнению с отрицательный контроль (фиг. 6А). Стимуляция клеток HaCaT с помощью ядерных клеток, индуцированных TNF-α / IFN-γ. транслокация p65 NF-κB и деградация IκB-α. NF-κB-зависимый анализ репортерного гена был использован для подтверждения ингибирующее действие RAME и Bay11-7082 на активацию NF-κB. РАМЭ и Bay11-7082 подавляли TNF-α / IFN-γ ядерную транслокация p65 NF-κB (рис.6Б). Как показано на фиг. 6C, TNF-α / IFN-γ стимулировали активность промотора NF-κB в клетках HaCaT был значительно снижен ингибиторами RAME и Bay11-7082. В Кроме того, стимуляция клеток HaCaT TNF-α / IFN-γ индуцировала экспрессия хемокинов и провоспалительных цитокинов. РАМЭ и Обработка Bay11-7082 подавляла экспрессию, стимулированную TNF-α / IFN-γ TARC, MDC, IL-6 и IL-8 (рис. 7A-D). Как показано на рис. 7, экспрессия хемокинов, стимулированных TNF-α / IFN-γ (TARC и MDC) и провоспалительные цитокины (IL-6 и IL-8) в HaCaT клеток значительно уменьшилось при обработке RAME и Ингибиторы Bay11-7082.
Рисунок 7Эффекты митоген-активированного белка ингибиторы киназ на экспрессию хемокинов и провоспалительные цитокины. Клетки HaCaT предварительно обрабатывали 20 мкг / мл RAME и 5 мкМ Bay11-7082, 10 мкМ SB203580, 10 мкМ PD98059 и 10 мкМ SP600125 в течение 1 часа с последующей инкубацией с 10 нг / мл TNF-α / IFN-γ в течение 18 часов. (А) TARC (B) MDC (C) уровни IL-8 и (D) IL-6 измеряли с помощью ELISA. Данные представлены как среднее значение ± стандартная ошибка среднего (n = 3).## P <0,01 по сравнению с нормальной контрольной группой, * P <0,05 и ** P <0,01 по сравнению с TNF-α / IFN-γ стимулированные клетки. РАМЭ, альбом рододендронов Блюм метанол извлекать; TNF-α, фактор некроза опухоли-α; IFN-γ, интерферон-γ; TARC, хемокин, регулируемый тимусом и активацией; MDC, полученный из макрофагов хемокин; ИЛ, интерлейкин. |
РАМЭ и ингибиторы ингибируют экспрессия хемокинов и цитокинов в TNF-α / IFN-γ-стимулированной Клетки HaCaT
Транслокация NF-κB в ядро была снижается в клетках HaCaT, обработанных RAME (рис.5). MAPK могут быть активированы TNF-α / IFN-γ, ключевой стимулятор воспалительного ответа кожи у кератиноциты, а также множество других типов клеток. В качестве представлен на рис. 4, RAME заметно снизил уровни экспрессии p-p38, p-ERK и p-JNK в TNF-α / IFN-γ-индуцированных клетках HaCaT по сравнению с необработанными клетками. Эти данные свидетельствуют о том, что RAME подавляет воспалительную реакцию. посредством частичной регуляции передачи сигналов NF-κB и MAPK. Чтобы оценить функция NF-κB и MAPK в производстве TNF-α / IFN-γ-индуцированные медиаторы воспаления, эффекты селективные ингибиторы NF-κB и MAPK на TNF-α / IFN-γ, индуцированные были исследованы продукция хемокинов и цитокинов.ELISA анализ выявили, что Bay11-7082 (ингибитор IκB-α) SP600125 (JNK ингибитор), PD98059 (ингибитор ERK) и SB203580 (ингибитор p38) заметно ингибирует экспрессию TARC, MDC, IL-6 и IL-8, соответственно. Кроме того, лечение RAME и ингибиторами было продемонстрировано, что заметно ингибирует TNF-α / IFN-γ-опосредованную экспрессию TARC (фиг. 7A), MDC (фиг. 7B), IL-8 (фиг. 7C) и IL-6 (фиг. 7D). Эти результаты показали, что RAME-опосредованная инактивация NF-κB p65, JNK, ERK и p38 MAPK может, по крайней мере частично, нести ответственность за подавление TARC, MDC, IL-8 и IL-6 в клетках HaCaT.Кроме того, TNF-α / IFN-γ стимуляция вызвала увеличение активации NF-κB и MAPKs, в то время как обработка RAME подавляла этот эффект.
Лечение РАМЭ подавляет активация JAK / STAT в клетках HaCaT
Путь JAK / STAT важен для иммунной система. Чтобы исследовать, привело ли лечение RAME к подавление TNF-α / IFN-γ-опосредованной активации JAK / STAT в HaCaT клеток, вестерн-блоттинг и иммуноцитохимия с использованием специфических Были использованы антитела (рис.8). Предварительная обработка RAME за 1 час до воздействия TNF-α / IFN-γ в Клетки HaCaT. TNF-α / IFN-γ увеличивал фосфорилирование JAK / STAT сигнализация на 1 ч. В результате было обнаружено, что RAME подавляет фосфорилирование пути JAK / STAT в TNF-α / IFN-γ-стимулированном Клетки HaCAT (рис. 8А). В Кроме того, было обнаружено, что лечение RAME снижает транслокацию STAT1 в клетках HaCaT с помощью иммуноцитохимии (рис. 8B). Эти результаты показали, что RAME ингибировал активацию JAK / STAT в клетках HaCaT.
Рисунок 8Влияние RAME на TNF-α / IFN-γ, индуцированное Активация NF-κB в клетках HaCaT.(A) Фосфорилирование JAK1, JAK2 и STAT1 анализировали с помощью вестерн-блоттинга. (B) Ячейка локализацию STAT1 определяли иммуноцитохимическим методом. С использованием DAPI (синий), ядра визуализировались и наблюдались при × 400 увеличение. Данные представлены как среднее значение ± стандартная ошибка среднее значение трех образцов. ## P <0,01 против отрицательного контроль; * P <0,05, ** P <0,01 и *** P <0,001 по сравнению с клетками, стимулированными TNF-α / IFN-γ. РАМЭ, Экстракт метанола Rhododendron album Blume; TNF-α, опухоль фактор некроза-α; IFN-γ, интерферон-γ; NF-κB, ядерный фактор-κB; JAK, киназа Януса; СТАТ, преобразователи сигналов и активаторы транскрипция; р-, фосфорилированный. |
Обсуждение
Народная медицина с использованием натуральных трав может помочь предотвращать и лечить несколько заболеваний, связанных с иммунной системой, включая БА, атопическое воспаление и аллергия (27). Настоящее исследование показало, что Лечение РАМЭ уменьшило выработку различных воспалительных и хемокины и цитокины, опосредованные аллергией в TNF-α / IFN-γ, индуцированных Клетки HaCaT за счет инактивации NF-κB и MAPK.
Регулирование выработки цитокинов играет центральную роль в патогенез аллергических расстройств.Cho et al (27) наблюдали повышенный уровень IL-6, Экспрессия ИЛ-8, ИЛ-10, ИЛ-23 и TGF-β у пациентов с БА по сравнению со здоровыми людьми. В настоящем исследовании RAME снижение экспрессии генов IL-8 и IL-6 в клетках HaCaT (рис. 3A-D). Эти данные позволяют предположить, что РАМЭ оказывает противовоспалительное действие, подавляя производство воспалительных хемокинов и цитокинов. Кроме того, хемокины и их рецепторы выполняют важную функцию при БА путем регулирование возникновения и обострения воспаления в ответ на аллергены (28,29).Джулиани и др. И Чен и др. (30,31) продемонстрировали, что сывороточные уровни MDC и TARC увеличиваются при AD, и это увеличение положительно коррелировал с тяжестью заболевания при АД. К лучшему Насколько нам известно, настоящее исследование является первым, демонстрирующим что RAME ингибирует TNF-α / IFN-γ-индуцированную экспрессию TARC и MDC в Клетки кератиноцитов HaCaT (рис. 2A-D).
NF-κB представляет собой часть важного провоспалительный сигнальный путь. NF-κB регулирует экспрессию некоторых видов астмы, воспалений, аллергии и иммуноассоциированных гены через деградацию IκB (30-32).Следовательно, RAME может иметь противоаллергический эффект, основанный на снижении уровня активированного NF-κB (Рис. 5A-C). Ингибитор IκB-α Bay11-7082 (33), что предотвращает фосфорилирование и активация p65 использовались, потому что специфические ингибиторные конструкции, которые снижают уровни p65 и IκB-α в Клетки HaCaT не идентифицированы (рис. 6А-С). Эффект специфического ингибитора NF-κB Bay11-7082 было исследовано и выявлено, что они значительно ингибируют активация NF-κB. Этот результат, по-видимому, связан с механизмами лежащие в основе противовоспалительных эффектов RAME, которые вызваны подавлением транскрипционной активности NF-κB.МАПК каскад — один из сигнальных путей, участвующих в иммунной ответы (24,34,35). Путь передачи сигналов MAPK регулирует множество клеточных процессов, включая экспрессию генов, гибель и выживание клеток, подвижность и пролиферация клеток (36,37). MAPK включают три основных подсемейства, а именно ERK p42 / p44, MAPK p38 и JNK. В Сообщалось, что ингибирование MAPK снижает синтез провоспалительные цитокины и их внутриклеточная передача сигналов пути и подавляют активацию NF-κB (38,39).TNF-α / IFN-γ стимуляция HaCaT клетки были подтверждены для активации трех основных модулей MAPK, в том числе ERK, JNK и p38. Лечение RAME, ингибитором p38 MAPK SB203580, ингибитор ERK PD98059 и ингибитор JNK SP600125 снижение TNF-α / IFN-γ экспрессии провоспалительных цитокинов (IL-6 и IL-8) и хемокинов (TARC и MDC) до исходного уровня ценности. Кроме того, эти ингибиторы и РАМЭ снижали провоспалительный цитокин (IL-6 и IL-8) и хемокин (TARC и MDC) на ~ 50% (рис.7). Эти результаты предполагают, что ингибирование JNK, ERK и p38 Лечение MAPK by RAME снижает выработку провоспалительных цитокины и хемокины в клетках HaCaT (рис. 4). Тот факт, что РАМЭ снижает TNF-α / IFN-γ-индуцированная экспрессия мРНК MAPK, MDC и TARC снова предположил, что РАМЭ обладает противоаллергическим и противовоспалительным действием. эффекты.
У млекопитающих путь JAK / STAT является основным сигнальный механизм для широкого спектра факторов роста и цитокины. Передача сигналов JAK / STAT является одним из множества плейотропных каскады, используемые для преобразования сигналов для развития у животных и люди.Результаты настоящего исследования показали, что RAME может иметь противоаллергический эффект, основанный на снижении активированного JAK / STAT (рис. 8A и B).
В заключение, настоящее исследование подтвердило, что RAME подавляет экспрессию хемокинов и цитокинов, связанных с AD в кератиноцитах, индуцированных TNF-α / IFN-γ. Эти эффекты считались связаны с подавлением активации NF-κB, эти экспериментальные результаты обеспечивают научную основу для использования РАМЭ лечить AD. Дополнительные эксперименты для тестирования in vitro эффекты RAME на AD в настоящее время продолжаются в нашем лаборатория.
Благодарности
Не применимо.
Банкноты
[1] Финансирование
Настоящее исследование было поддержано Bio & Программа развития медицинских технологий национального исследования Фонд и финансируется правительством Кореи (MSIT; грант No. NRF-2016K1A1A8A01939075) и Корейский научно-исследовательский институт Инициативная программа исследований в области бионауки и биотехнологии Республика Корея (грант № KGM1221814).
[2] Наличие данных и материалов
Проанализированные наборы данных, полученные в ходе исследования. доступны у соответствующего автора по разумной запрос.
[3] Авторские взносы
JWP проанализировал данные и написал рукопись. HSL и YRL провели эксперименты in vitro. JHP, OKK, JHK, IP и PY подготовили альбом Rhododendron Blume, а проанализировал и отредактировал рукопись. СК, СРО и КСА разработали изучил и отредактировал рукопись. Все авторы критически пересмотрели статью и согласились с окончательной версией рукопись.
[4] Этика согласование и согласие на участие
Не применимо.
[5] Согласие на публикация
Не применимо.
[6] Конкуренция проценты
Авторы заявляют, что у них нет конкурирующих интересы.
Список литературы
1 | Brandt EB и Sivaprasad U: цитокины Th3 и атопический дерматит. J. Clin Cell Immunol. 2: pii. 1102011. Просмотр статьи: Google Scholar: PubMed / NCBI |
2 | Абрамовиц В. Атопический дерматит.J Am Acad Дерматол. 53 (1 приложение 1): S86 – S93. 2005. Просмотр статьи: Google Scholar: PubMed / NCBI |
3 | Португалия-Коэн М., Хорев Л., Раффер С., Schlippe G, Voss W, Ma’or Z, Oron M, Soroka Y, Frušić-Zlotkin M, Милнер Ю. и Коэн Р.: Количественная оценка неинвазивных биомаркеров кожи псориаза и атопического дерматита: цитокины, антиоксиданты и псориатическая аутофлуоресценция кожи. Biomed Pharmacother. 66: 293–299. 2012. Просмотр статьи: Google Scholar: PubMed / NCBI |
4 | Хоффьян С. и Стеммлер С. О роли комплекс эпидермальной дифференцировки при вульгарном ихтиозе, атопический дерматит и псориаз.Br J Dermatol. 157: 441–449. 2007 г. Просмотр статьи: Google Scholar: PubMed / NCBI |
5 | Накацудзи Т., Чен Т.Х., Два часа ночи, Чун К.А., Нарала С., Геха Р.С., Хата Т.Р. и Галло Р.Л.: золотистый стафилококк. использует дефекты эпидермального барьера при атопическом дерматите, чтобы вызвать экспрессия цитокинов. J Invest Dermatol. 136: 2192–2200. 2016 г. Просмотр статьи: Google Scholar: PubMed / NCBI |
6 | Леунг Д.Ю., Богуневич М., Хауэлл, доктор медицины, Номура Я и Хамид QA: Новые взгляды на атопический дерматит.J Clin Invest. 113: 651–657. 2004. Просмотр Статья: Google Scholar: PubMed / NCBI |
7 | Ян Ай Джей, Ли ДЮ и Шин Х.М.: Тормозящие влияние валенсена на развитие атопического дерматита поражения кожи у мышей nc / nga. Evid Based Complement Alternat Med. 2016: 93708932016. Просмотр статьи: Google Scholar: PubMed / NCBI |
8 | Злотник А и Йоши О: Хемокины: новый системы классификации и их роль в иммунитете.Иммунитет. 12: 121–127. 2000. Просмотр статьи: Google Scholar: PubMed / NCBI |
9 | Кан Г.Дж., Хан С.К., Йи Э.Дж., Кан Г.К. и Ю ES: Ингибирующее действие экстракта преждевременного цитрусового уншиу на атопический дерматит in vitro и in vivo. Toxicol Res. 27: 173–180. 2011. Просмотр статьи: Google Scholar |
10 | Вестергаард С, Йонеяма Х, Мурай М, Накамура К., Тамаки К., Терашима Ю., Имаи Т., Йоши О, Иримура Т., Mizutani H и Matsushima K: перепроизводство специфичных для Th3 хемокины у мышей NC / Nga, проявляющих атопический дерматитоподобный поражения.J Clin Invest. 104: 1097–1105. 1999. Просмотр статьи: Google Scholar: PubMed / NCBI |
11 | Хорикава Т., Накаяма Т., Хикита И., Ямада H, Fujisawa R, Bito T, Harada S, Fukunaga A, Chantry D, Gray PW и др. al: IFN-гамма-индуцируемая экспрессия тимуса и регулируемый активацией хемокин / CCL17 и происходящие из макрофагов хемокин / CCL22 в эпидермальных кератиноцитах и их роль в атопический дерматит. Int Immunol. 14: 767–773. 2002. Просмотр статьи: Google Scholar: PubMed / NCBI |
12 | Себастьяни С., Албанези К., Де ПО, Пудду П., Cavani A и Girolomoni G: роль хемокинов при аллергических заболеваниях. контактный дерматит.Arch Dermatol Res. 293: 552–559. 2002 г. Просмотр статьи: Google Scholar: PubMed / NCBI |
13 | Shimada Y, Takehara K и Sato S: оба Th3 и хемокины Th2 (TARC / CCL17, MDC / CCL22 и Mig / CXCL9) являются повышен в сыворотках крови пациентов с атопическим дерматитом. J Dermatol Sci. 34: 201–208. 2004. Просмотр статьи: Google Scholar: PubMed / NCBI |
14 | Кимура М, Хайса М, Уэцука Х, Такаока М, Окава Т., Кавасима Р., Ямацудзи Т., Гундуз М., Канеда Ю., Танака Н. и Naomoto Y: TNF в сочетании с IFN-альфа ускоряет NF-kappaB-опосредованный апоптоз за счет увеличения экспрессии Fas в раковых клетках толстой кишки.Смерть клетки отличается. 10: 718–728. 2003 г. Просмотр статьи: Google Scholar: PubMed / NCBI |
15 | Rath PC: взаимосвязь между конститутивными ядерный фактор-каппаВ (NF-каппаВ) и ингибитор каппаВ-альфа (IkappaB-alpha) у человека Беркитта, чувствительного к интерферон-альфа линия клеток лимфомы. Biochim Biophys Acta. 1741: 253–263. 2005 г. Просмотр статьи: Google Scholar: PubMed / NCBI |
16 | Baeuerle PA и Baichwal VR: NF-kappa B as частая мишень для иммуносупрессивных и противовоспалительных молекулы.Adv Immunol. 65: 111–137. 1997. Просмотр статьи: Google Scholar: PubMed / NCBI |
17 | Ортис Ф., Пирот П., Наамане Н., Крейнс А.Ю., Расшерт Дж., Мур Ф., Театр Э, Верхаге С., Магнуссон Н. Э., Chariot A и др.: Индукция ядерного фактора-каппаB и его нижестоящие гены TNF-альфа и IL-1beta играют проапоптотическую роль в бета-клетках поджелудочной железы. Диабетология. 51: 1213–1225. 2008 г. Просмотр статьи: Google Scholar: PubMed / NCBI |
18 | Цао Чж, Инь В.Д., Чжэн Цюй, Фэн С.Л., Сюй Г.Л. и Zhang KQ: каспаза-3 участвует в опосредованных IFN-γ и TNF-α Апоптоз клеток MIN6 по пути NF-κB / Bcl-2.Cell Biochem Biophys. 67: 1239–1248. 2013. Просмотр статьи: Google Scholar |
19 | Али С., Нисар М., Кайсар М., Хан А. и Хан AA: Оценка цитотоксического потенциала нового пентациклического тритерпен из коры стебля Rhododendron arboreum. Pharm Biol. 55: 1927–1930. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI |
20 | Ли Ф.Р., Юй FX, Яо С.Т., Си Й.Х., Чжан В. и Gao LL: Гиперин, извлеченный из листьев маньчжурского рододендрона, вызывает апоптоз в клетках рака эндометрия человека через митохондриальные путь.Азиатский Pac J Cancer Prev. 13: 3653–3656. 2012. Просмотр статьи: Google Scholar: PubMed / NCBI |
21 | Лю Ю.Л., Линь Л.С., Тунг Ю.Т., Хо С.Т., Чен Ю.Л., Lin CC и Wu JH: Экстракт листьев Rhododendron oldhamii улучшает жирность печеночный синдром за счет увеличения окисления липидов и уменьшения путь липогенеза у мышей. Int J Med Sci. 14: 862–870. 2017 г. Просмотр статьи: Google Scholar: |
22 | Пак Дж. У., Квон ОК, Ким Дж. Х., О СР, Ким Дж. Х., Пайк Дж. Х., Марвото Б., Виджхати Р., Джуниарти Ф., Ираван Д. и Ан К. С.: Альбом рододендрона Blume подавляет экспрессию iNOS и COX-2 в LPS-стимулированный RAW264.7 клеток через подавление NF- κB сигнализация. Int J Mol Med. 35: 987–994. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI |
23 | Ли Дж. У., Пак Дж. В., Квон ОК, Ли ХД, Чжон HG, Kim JH, Oh SR и Ahn KS: NPS2143 ингибирует MUC5AC и провоспалительные медиаторы в экстракте сигаретного дыма (CSE) -стимулируемые эпителиальные клетки дыхательных путей человека. Воспаление. 40: 184–194. 2017. Просмотр статьи: Google Scholar |
24 | Ко Дж. У., Пак Дж. В., Шин Н. Р., Ким Дж. Х., Чо Ю. К., Shin DH, Kim JC, Lee IC, Oh SR, Ahn KS и Shin IS: оксид меди наночастица вызывает воспалительную реакцию и выработку слизи за счет Передача сигналов MAPK в эпителиальных клетках бронхов человека.Environ Toxicol Pharmacol. 43: 21–26. 2016. Просмотреть статью: Google Scholar: PubMed / NCBI |
25 | Ливак К.Дж. и Шмитген Т.Д.: Анализ данные об относительной экспрессии генов с использованием количественной ПЦР в реальном времени и метод 2 (-Delta Delta C (T)). Методы. 25: 402–408. 2001 г. Просмотр статьи: Google Scholar |
26 | Шибата С., Маэда С., Кондо Н., Чимура Н., Иноуэ А и Фуката Т. Идентификация сигнального пути TNF-α-индуцированная транскрипция CCL17 / TARC в кератиноцитах собаки клеточная линия.Vet Immunol Immunopathol. 139: 90–98. 2011. Просмотр статьи: Google Scholar |
27 | Чо БО, Че Д.Н., Инь ХХ, Шин Джи и Чан SI: Экстракт листьев лотоса Diospyros и стеблей грейпфрута синергетически облегчает поражение кожи, подобное атопическому дерматиту, при мышей, подавляя проникновение тучных клеток в кожные поражения. Biomed Pharmacother. 89: 819–826. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI |
28 | Homey B, Steinhoff M, Ruzicka T и Leung DY: Цитокины и хемокины организуют атопическое воспаление кожи.J Allergy Clin Immunol. 118: 178–189. 2006. Просмотр статьи: Google Scholar: PubMed / NCBI |
29 | Недошитко Б, Соколовская-Войдыло М, Ruckemann-Dziurdzińska K, Roszkiewicz J и Nowicki RJ: Chemokines и сеть цитокинов в патогенезе воспалительного процесса кожи заболевания: атопический дерматит, псориаз и мастоцитоз кожи. Постэпы Дерматол Алергол. 31: 84–91. 2014. Просмотр статьи: Google Scholar: PubMed / NCBI |
30 | Джулиани К, Наполитано Дж, Буччи I, Монтани V и Монако F: фактор транскрипции Nf-κB: роль в патогенез воспалительных, аутоиммунных и опухолевых заболеваний и значение терапии.Clin Ter. 152: 249–253. 2001 г. на итальянском языке. PubMed / NCBI |
31 | Чен Й, Сиань Й, Лай З, Лу С, Чан Вай и Lin ZX: противовоспалительное и противоаллергическое действие и лежащие в основе механизмы экстракта Хуан-Лянь-Цзе-Ду: значение для атопического лечение дерматита. J Ethnopharmacol. 185: 41–52. 2016. Просмотреть статью: Google Scholar: PubMed / NCBI |
32 | Пак JW, Ким YJ, Шин ИС, Квон ОК, Хонг JM, Shin NR, Oh SR, Ha UH, Kim JH и Ahn KS: секреция типа III система Pseudomonas aeruginosa влияет на матричную металлопротеиназу 12 (MMP-12) и MMP-13 Экспрессия посредством передачи сигналов ядерного фактора κB в эпителиальных клетках карциномы человека и мышиной модели пневмонии.J Заразить Dis. 214: 962–969. 2016. Просмотреть статью: Google Scholar: PubMed / NCBI |
33 | Пак Дж. У., Шин И.С., Ха УХ, О СР, Ким Дж. Х и Ан К.С.: Патофизиологические изменения, вызванные Pseudomonas aeruginosa вовлечены в активацию MMP-12 и MMP-13 в эпителиальных клетках карциномы человека и мышиной модели пневмонии. Заражение иммунной. 83: 4791–4799. 2015. Просмотр статьи: Google Scholar: PubMed / NCBI |
34 | Kim HH, Kim SW, Kim DS, Oh HM, Rho MC и Ким Ш.: Vigna angularis подавляет аллергические реакции, опосредованные тучными клетками. воспаление.Int J Mol Med. 32: 736–742. 2013. Просмотр статьи: Google Scholar: PubMed / NCBI |
35 | Пак Дж. У., Ли IC, Шин Н. Р., Чон СМ, Квон ОК, Ко Дж. У., Ким Дж. Си, О СР, Шин И.С. и Ан К.С.: Оксид меди наночастицы усугубляют воспаление дыхательных путей и выработку слизи в астматические мыши с помощью передачи сигналов MAPK. Нанотоксикология. 10: 445–452. 2016. Просмотр статьи: Google Scholar |
36 | Jeong YH, Oh YC, Cho WK, Lee B и Ma JY: Противовоспалительное действие этанольного экстракта травы меландрии через ингибирование сигнальных путей NF-κB и MAPK и индукция HO-1 в RAW 264.7 клеток и первичные макрофаги мыши. Молекулы. 21: pii. E8182016. Просмотр статьи: Google Scholar |
37 | Чжэн Г., Шен З, Чен Х, Лю Дж, Цзян К., Fan L, Jia L и Shao J: метапристон подавляет немелкие клетки распространение и метастазирование рака легких путем модуляции Сигнальный путь RAS / RAF / MEK / MAPK. Biomed Pharmacother. 90: 437–445. 2017. Просмотр статьи: Google Scholar: PubMed / NCBI |
38 | Чжао С., Сунь И, Ли Х, Ван Дж, Ян Л., Чжан Z, Wang D, Dai J, He J и Wang S: скутелларин подавляет RANKL-опосредованный остеокластогенез и индуцированный частицами титана остеолиз посредством подавления сигнального пути NF-κB и MAPK.Int Иммунофармакол. 40: 458–465. 2016. Просмотреть статью: Google Scholar: PubMed / NCBI |
39 | Пак Дж. У., Квон ОК, Юниато П., Марвото Б., Lee J, Oh SR, Kim JH и Ahn KS: улучшение вызванного LPS воспалительный ответ с использованием метанольного экстракта лагерстроемии ovalifolia для подавления активации NF-κB в RAW264.7 макрофаги. Int J Mol Med. 38: 482–490. 2016. Просмотреть статью: Google Scholar: PubMed / NCBI |
Интерфероновая резистентность новых вариантов SARS-CoV-2
Abstract
Появление вариантов SARS-CoV-2 с повышенной трансмиссивностью, патогенезом и устойчивостью к вакцинам представляет собой неотложную проблему для сдерживания пандемии COVID-19.Хотя спайковые мутации, повышающие инфекционность вируса, могут способствовать появлению этих новых вариантов, исследования, документально подтверждающие критическую роль ответов интерферона в раннем контроле инфекции SARS-CoV-2, в сочетании с наличием вирусных генов, ограничивающих эти ответы, предполагают что интерфероны также могут влиять на развитие SARS-CoV-2. Здесь мы сравнили эффективность 17 различных человеческих интерферонов против 5 вирусных линий, отобранных в ходе глобальной вспышки, которые включали предковые и новые варианты.Наши данные выявили повышенную резистентность к интерферону у появляющихся вариантов SARS-CoV-2, что указывает на то, что уклонение от врожденного иммунитета является важной движущей силой для эволюции SARS-CoV-2. Эти результаты имеют значение для увеличения летальности появляющихся вариантов и выделяют подтипы интерферона, которые могут быть наиболее успешными при лечении ранних инфекций.
Геном человека кодирует разнообразный набор противовирусных интерферонов (IFN). К ним относятся IFN типа I (IFN-Is), такие как 12 подтипов IFNα, IFNβ и IFNω, которые передают сигнал через повсеместно распространенный рецептор IFNAR, и IFN типа III (IFN-III), такие как IFNλ1, IFNλ2 и IFNλ3, которые передают сигнал через более ограниченный рецептор IFNλR, который присутствует в эпителиальных клетках легких 1 .Разнообразие IFN может быть обусловлено эволюционной гонкой вооружений, позволяющей хозяину противодействовать различным вирусным патогенам 2 . Например, подтипы IFNα демонстрируют> 78% идентичности аминокислотной последовательности, но IFNα14, IFNα8 и IFNα6 наиболее эффективно ингибируют ВИЧ-1 in vitro и in vivo 3-5 , тогда как IFNα5 наиболее эффективно ингибирует h4N2 гриппа в Культуры эксплантатов легких 6 . Удивительно, хотя SARS-CoV-2 был чувствителен к IFNα2, IFNβ и IFNλ 7–9 , а клинические испытания IFNα2 и IFNβ продемонстрировали многообещающие результаты против COVID-19 10–12 , прямое сравнение нескольких IFN- Is и IFN-III против различных изолятов SARS-CoV-2 еще не проводились.
Результаты
Настоящее исследование было предпринято для определения того, какие интерфероны лучше всего ингибируют SARS-CoV-2. Мы отобрали 5 изолятов из известных линий 13 во время пандемии (рис. 1, дополнительная таблица 1). USA-WA1 / 2020 — это стандартный штамм, используемый во многих исследованиях in vitro, и in vivo, SARS-CoV-2, и принадлежит к линии A 13 . Он был изолирован от первого пациента с COVID-19 в США, который имел прямую эпидемиологическую связь с Ухань, Китай, где вирус был впервые обнаружен 14 .Напротив, последующие волны инфекции из Азии в Европу 15 были связаны с появлением мутации D614G 16 . Штаммы D614G + в линии B распространяются с невероятной скоростью, вероятно, из-за его повышенной трансмиссивности 17,18 . Он накопил дополнительные мутации в Италии в виде линии B.1, которые затем спровоцировали серьезную вспышку в Нью-Йорке 19 . Совсем недавно линия B.1.1.7 приобрела мутацию N501Y, которая связана с повышенной трансмиссивностью в Соединенном Королевстве 13 .Линия B.1.351 была впервые обнаружена в Южной Африке и приобрела дополнительную мутацию E484K, которая связана с устойчивостью к нейтрализующим антителам 20,21 . И B.1.1.7, и B.1.351 в настоящее время зарегистрированы во многих странах, и растет беспокойство по поводу того, что они могут стать доминирующими. 22 . Репрезентативные изоляты SARS-CoV-2 из линий B, B.1, B.1.1.7 и B.1.351 были получены из BEI Resources (дополнительная таблица 1) и амплифицированы один раз в линии эпителиальных клеток альвеолярного типа II, A549, которая мы стабильно трансдуцировали рецептором ACE2 (A549-ACE2) (дополнительный рис.1а).
Рисунок 1 | Выбор штаммов SARS-CoV-2 для исследований чувствительности к интерферону.( a ) Глобальное распространение клад SARS-CoV-2. GISAID.org построил график соотношения депонированных последовательностей в обозначенных кладах в зависимости от дат сбора. Выбранные пять изолятов отмечены цветными точками. ( b ) Штаммы SARS-CoV-2, выбранные для этого исследования, включали представителей линий A, B, B.1, B.1.351 и B.1.1.7 (дополнительная таблица 1). Изоляты линии B кодируют мутацию D614G, связанную с повышенной трансмиссивностью.* Аминокислотные мутации относились к эталонной последовательности hCOV-19 / Wuhan / WIV04 / 2019.
Клетки A549-ACE2 предварительно инкубировали с 17 рекомбинантными IFN (PBL Assay Science) в течение ночи параллельно и в трех повторностях, затем инфицировали ненасыщающей дозой вируса в течение 2 часов (дополнительный рисунок 1b). Мы нормализовали IFN на основе молярных концентраций, как и в нашей предыдущей работе с ВИЧ-1 3,23 . Для быстрой и надежной оценки противовирусной активности против живых изолятов SARS-CoV-2 мы использовали метод количественной ПЦР (рис.2а). Первоначальное исследование титрования дозы показало, что концентрация 2 пМ максимально различает противовирусную активность IFNβ и IFNλ1 (дополнительный рис. 1c), и поэтому использовалась для скрининга противовирусных IFN. В отсутствие IFN все 5 изолятов достигли титров ~ 10 4 -10 6 копий на 5 мкл введенного экстракта РНК (рис. 2). Используя абсолютное число копий (рис. 2) и значения, нормализованные до фиктивного, равного 100% (дополнительный рис. 2), 17 IFN продемонстрировали диапазон противовирусной активности против SARS-CoV-2.Три подтипа IFN-λ проявляли от нулевой до очень слабой (<2-кратной) противовирусной активности по сравнению с большинством IFN-Is (рис. 2 и дополнительный рис. 2, синие столбцы). И это несмотря на то, что анализ показал устойчивый динамический диапазон, при этом некоторые ИФН ингибируют USA-WA1 / 2020 в> 2500 раз и ниже обнаруживаемых уровней (рис. 2а). Эффективность ИФН в отношении 5 изолятов коррелировала друг с другом (дополнительный рисунок 3), и аналогичный порядок ранжирования противовирусной активности ИФН наблюдали для изолятов D614G + (рисунок 2b, дополнительный рисунок.2). В целом, IFNα8, IFNβ и IFNω были наиболее эффективными, за ними следовали IFNα5, IFNα17 и IFNα14 (рис. 2c).
Рисунок 2 | Чувствительность штаммов SARS-CoV-2 к интерферонам IFN-I и IFN-III.( a ) Противовирусный анализ с использованием рекомбинантных IFN (2 пМ) в клетках A549-ACE2. Красная линия соответствует пределу обнаружения КПЦР (<90 копий / реакция). (b ) Число вирусных копий в изолятах D614G +, показывающее аналогичный порядок ранжирования IFN от наименее до наиболее сильного. ( c ) Среднее ингибирование сворачивания относительно фиктивного для линий B, B.1 показаны изоляты B.1.351 и B.1.1.7. Наиболее эффективные IFN показаны сверху вниз. Для всех панелей столбцы и планки погрешностей соответствуют средним значениям и стандартным отклонениям.
Мы сообщили, что ингибирование ВИЧ-1 подтипами IFNα коррелирует с сигнальной способностью IFNAR и аффинностью связывания с субъединицей IFNAR2 3,23 . Способность передачи сигналов IFNAR, измеренная на линии IFN-чувствительных репортерных клеток (клетки iLite; Euro Diagnostics), коррелировала с противовирусной эффективностью подтипов IFNα в отношении линий SARS-CoV-2 A и B, но не B.1, B.1.351 или B.1.1.7 (рис. 3а). Интересно, что ингибирование SARS-CoV-2 подтипом IFNα не коррелировало со сродством связывания IFNAR2 (рис. 3b) 24 , как измерено с помощью поверхностного плазмонного резонанса группой Шрайбера 24 . Кроме того, корреляция между ингибированием SARS-CoV-2 и ВИЧ-1 3 была в лучшем случае слабой (рис. 3c). Эти данные свидетельствуют о том, что IFN-опосредованный контроль изолятов SARS-CoV-2 может качественно отличаться от контроля ВИЧ-1.
Рисунок 3 | Корреляция между ингибированием SARS-CoV-2 и биологическими свойствами подтипов IFNα.Log-трансформированные значения ингибирования IFN по сравнению с имитацией для 5 различных штаммов SARS-CoV-2 сравнивали с ранее опубликованными значениями для ( a ) эффективных концентраций 50% в анализе iLite, репортерной клеточной линии, кодирующей чувствительный к IFN элемент ответа ISG15 , связанный с люциферазой светлячка 23 ; (b ) Аффинность связывания субъединицы IFNAR2, измеренная с помощью поверхностного плазмонного резонанса группой Шрайбера 24 ; и ( c ) значения ингибирования ВИЧ-1, основанные на% ингибирования p24 + лимфоцитов кишечника ВИЧ-1 по сравнению с ложными, как измерено проточной цитометрией 3 .Каждая точка соответствует подтипу IFNα. Линейную регрессию выполняли с использованием GraphPad Prism 8. Значимые корреляции ( p <0,05) были выделены красной линией наилучшего соответствия; у тех, которые были в тренде ( p <0,1), была серая пунктирная линия наилучшего соответствия.
Мы создали тепловую карту для визуализации противовирусной активности различных IFN против 5 изолятов и наблюдали заметные различия в чувствительности IFN (рис. 4a). Попарный анализ противовирусной активности изолятов, собранных в начале (январь 2020 г.) и позднее (март-декабрь 2020 г.) во время пандемии, был проведен в отношении 14 IFN-Is (IFN-III не были включены из-за низкого ингибирования, рис.2). Общая чувствительность к IFN-I изолятов USA-WA1 / 2020 и Germany / BavPat1 / 2020 существенно не отличалась друг от друга (рис. 4b). Напротив, по сравнению с Germany / BavPat1 / 2020, мы наблюдали от 17 до 122-кратной устойчивости к IFN-I возникающих вариантов SARS-CoV-2 (рис. 4c), при этом штамм B.1.1.7 демонстрировал самый высокий уровень IFN-1. Я сопротивление. Уровень резистентности к интерферону был более поразительным по сравнению с USA-WA1 / 2020, где появляющиеся варианты SARS-CoV-2 демонстрировали в 25-322 раза более высокую резистентность к IFN-I (дополнительный рис.4а).
Рисунок 4 | Повышенная резистентность к интерферону возникающих вариантов SARS-CoV-2.( a ) Тепловая карта кратковременного ингибирования репрезентативных штаммов отмеченных линий. Цвета оценивали по логарифмической шкале от наивысшего ингибирования (желтый) до отсутствия ингибирования (черный). Сравнение чувствительности к IFN-I между изолятами линий A и B ( b ); и ( c ) линия B по сравнению с B.1, B.1.351 и B.1.1.7. Средние значения кратного ингибирования относительно ложного сравнивали попарно для 14 IFN-Is.В ( c ) были отмечены средние значения ингибирования сворачивания. Различия оценивали с помощью непараметрического двустороннего критерия ранговых знаков Уилкоксона для согласованных пар. NS, не имеет значения; ****, п. <0,0001. ( d ) Титрование дозы IFNβ и IFNλ1 по линии B (Германия / BavPat1 / 2020) по сравнению с изолятами B.1.1.7 и B.1.351. Помимо USA / CA_CDC_5574 / 2020, мы также оценили второй изолят B.1.1.7 из Соединенного Королевства (UK), England / 204820464/2020. Клетки A549-ACE2 предварительно обрабатывали серийными 10-кратными разведениями IFN в течение 18 часов в трех экземплярах, а затем инфицировали SARS-CoV-2.Супернатанты собирали через 24 часа, количество копий SARS-CoV-2 N1 определяли с помощью кПЦР, а затем нормализовали по ложному результату как 100%. Для интерполяции 50% ингибирующих концентраций (зеленые пунктирные линии) использовали нелинейные кривые регрессии наилучшего соответствия средних нормализованных уровней инфекции.
Вышеуказанные эксперименты позволили провести одновременный анализ 17 IFN против нескольких изолятов SARS-CoV-2, но не предоставили информации о том, как разные дозы IFN-I влияют на репликацию вируса. Также остается неясным, были ли появляющиеся варианты устойчивыми к IFN-III.Поэтому мы титровали сильный (IFNβ; от 0,002 до 200 пМ) и слабый (IFNλ1; от 0,02 до 2000 пМ) интерферон против изолятов линий B, B.1, B.1.1.7 и B.1.351 (рис. 4d и дополнительные Рис. 4б). Мы включили дополнительный штамм B.1.1.7, hCov-19 / England / 204820464/2020 (дополнительная таблица 1). 50% ингибирующие концентрации (IC 50 ) вариантов B.1.1.7 были в 4,3-8,3 раза выше для IFNβ и в 3,0-3,5 раза выше для IFNλ1, чем у изолята линии B (рис. 4d), тогда как B. Показан 1 изолят 2.IC 50 для IFNλ1 и IFNβ в 6 и 5,5 раз выше соответственно (дополнительный рис. 4b). Интересно, что максимальное ингибирование не было достигнуто ни с IFNβ, ни с IFNλ1 против варианта B.1.1.7, достигая плато на уровнях в 15-20 раз выше, чем у изолята предковой линии B (фиг. 4d). В отдельном эксперименте вариант B.1.351 также был более устойчив к IFNβ (> 500 раз) и IFNλ1 (26 раз) по сравнению с изолятом линии B (фиг. 4d). Эти данные подтверждают, что B.1, B.1.1.7 и B.1.351 изолят эволюционировал, чтобы противостоять ответу IFN-I и IFN-III.
Обсуждение
Многочисленные исследования, проведенные многими лабораториями, подчеркнули важность IFNs в борьбе с SARS-CoV-2. Здесь мы демонстрируем продолжающуюся эволюцию SARS-CoV-2, чтобы избежать ответов IFN, и идентифицировать IFN с наивысшей противовирусной активностью. IFNλ первоначально показал себя многообещающим в качестве противовирусного средства, которое может уменьшать воспаление 25 , но недавно был связан с вирусной патологией легких 26 .Наши данные показывают, что для достижения аналогичного противовирусного эффекта in vivo , как и IFN-Is, могут потребоваться более высокие дозы IFN-λ. Распыленный IFNβ показал потенциал в качестве терапевтического средства против COVID-19 11 , и наши данные подтверждают, что IFNβ является высокоэффективным противовирусным средством против SARS-CoV-2. Однако IFNβ также был связан с патогенетическими исходами при хроническом инфицировании слизистых оболочек ВИЧ-1 23 , LCMV 27 у мышей и при позднем введении мышам SARS-CoV-1 и MERS-CoV 28,29 .Напротив, IFNα8 изменял в 3 раза меньше генов в первичных лимфоцитах слизистой оболочки, чем IFNβ 23 , но показал аналогичную эффективность против SARS-CoV-2, как IFNβ. IFNα8 также проявлял высокую противовирусную активность против ВИЧ-1 3 , повышая его потенциал для лечения обоих пандемических вирусов. Примечательно, что IFNα8 оказался необычным, поскольку противовирусная активность подтипов IFNα против SARS-CoV-2 и ВИЧ-1 не сильно коррелировала. IFNα6 сильно ограничивал ВИЧ-1 3,4 , но был одним из самых слабых подтипов IFNα против SARS-CoV-2.И наоборот, IFNα5 сильно ингибирует SARS-CoV-2, но слабо ингибирует HIV-1 3 . Наши данные подтверждают теорию о том, что различные IFN, возможно, эволюционировали, чтобы ограничить отдельные семейства вирусов 2,23 . Механизмы, лежащие в основе этих качественных различий, остаются неясными. В то время как передача сигналов IFNAR способствует противовирусной активности 3,4,24 , различные IFN могут обладать различными способностями мобилизовать противовирусные эффекторы в определенных типах клеток. Сравнение интерферомов, индуцируемых отдельными IFN в эпителиальных клетках легких, может помочь раскрыть противовирусные механизмы, ответственные за различные эффекты.
Наши данные выявили тревожную тенденцию к появлению вариантов SARS-CoV-2, которые сопротивляются противовирусному ответу IFN. До этой работы возникновение и фиксация вариантов было связано с повышенной вирусной инфекционностью из-за мутаций в белке Spike 13,16–18 . Однако предыдущие исследования инфекции ВИЧ-1 показали, что IFN также могут влиять на эволюцию пандемических вирусов 30,31 . Фактически, у инфицированных SARS-CoV-2 людей с генетическими дефектами передачи сигналов IFN 32 или IFN-реактивных аутоантител 33 был повышенный риск развития тяжелой формы COVID-19.Поскольку интерфероны имеют решающее значение для контроля ранних уровней вирусной инфекции, устойчивые к интерферону варианты SARS-CoV-2 могут вызывать более высокую вирусную нагрузку, что, в свою очередь, может способствовать передаче и / или усугублять патогенез. В соответствии с этой гипотезой тревожные предварительные отчеты связывают B.1.1.7 с повышенной вирусной нагрузкой 34 и риском смерти 35–37 . Помимо Spike, появляющиеся варианты обнаруживали мутации в нуклеокапсиде, мембранных и неструктурных белках NSP3, NSP6 и NSP12 (дополнительная таблица 1).Было показано, что эти вирусные белки противодействуют передаче сигналов IFN в клетках 38-40 . Будет важно определить вирусные мутации, вызывающие резистентность к IFN-I в новых вариантах, лежащие в основе молекулярные механизмы и его последствия для патогенеза COVID-19.
В целом, текущее исследование предполагает роль врожденного иммунного ответа в развитии SARS-CoV-2, что может иметь практическое значение для терапии на основе интерферона. Наши результаты подтверждают важность продолжения полногеномного надзора за SARS-CoV-2 и оценки появляющихся вариантов не только с точки зрения устойчивости к нейтрализующим антителам, вызываемым вакциной, но и с точки зрения уклонения от ответа хозяина на интерферон.
Материалы и методы
Клеточные линии
Клетки A549 были получены из Американской коллекции типовых культур (АТСС) и культивированы в полной среде, содержащей среду Хэма F-12 (Corning), 10% фетальную бычью сыворотку (Atlanta Biologicals), 1 % пенициллин / стрептомицин / глутамин (Corning) и поддерживается при 37 ° C 5% CO 2 . Клетки A549 трансдуцировали с помощью оптимизированного по кодонам человеческого ACE2 (Genscript), клонированного в pBABE-puro 41 (Addgene). Для создания стабильной клеточной линии A549-ACE2 10 7 HEK293T (ATCC) клеток в колбах T-175 временно котрансфицировали 60 мкг смеси pBABE-puro-ACE2, pUMVC и pCMV-VSV-G при Соотношение 10: 9: 1 с использованием кальций-фосфатного метода 42 .Через 48 часов после трансфекции супернатант собирали, центрифугировали при 1000 × g в течение 5 минут и пропускали через шприцевой фильтр 0,45 мкм для удаления клеточного дебриса. Отфильтрованный вирус смешивали со свежей средой (30% об. / Об.), Которая включала полибрен (Sigma) в конечной концентрации 6 мкг / мл. Смесь вирусов добавляли в 6-луночные планшеты с 5 × 10 5 клеток A549 / лунку и среду меняли еще раз через 12 часов. Трансдуцированные клетки отбирали в пуромицине 0,5 мкг / мл в течение 72 часов, и экспрессию ACE2 подтверждали проточной цитометрией, вестерн-блоттингом и чувствительностью к псевдовирионам HIV-1ΔEnv / SARS-CoV-2 Spike.
Изоляты вируса
Все эксперименты с живым вирусом SARS-CoV-2 были выполнены в учреждении уровня биобезопасности-3 (BSL3) с очищающими воздух респираторами с электроприводом в медицинском кампусе Университета Колорадо в Аншуце. Запасы SARS-CoV-2 из BEI Resources (дополнительная таблица 1) имели сопоставимые титры> 10 6 TCID 50 / мл (дополнительный рисунок 1a), за исключением штаммов B.1.1.7 (CA_CDC_5574 / 2020 и England / 204820464/2020). Содержимое всего флакона (~ 0,5 мл) инокулировали в 3 колбы Т-75, содержащих 3 × 10 6 клеток A549-ACE2, за исключением B.1.1.7, который засевали в 1 колбу Т-75. После культивирования в течение 72 часов супернатанты собирали и центрифугировали при 2700 × г в течение 5 минут для удаления остатков клеток и замораживали при -80 ° C. Штоки, амплифицированные A549, были титрованы в соответствии с предложенным форматом анализа (дополнительный рис. 1b, рис. 2a). Вкратце, 2,5 × 10 4 клеток A549-ACE2 высевали на лунку в 48-луночный планшет в течение ночи. На следующий день клетки инфицировали 300, 30, 3, 0,3, 0,03 и 0,003 мкл (серийное 10-кратное разведение) запаса амплифицированного вируса в 300 мкл конечного объема среды в течение 2 часов.Вирус дважды промывали PBS и добавляли 500 мкл полной среды с соответствующими концентрациями IFN. Через 24 ч супернатанты собирали и остатки клеток удаляли центрифугированием при 3200 × g в течение 5 минут.
SARS-CoV-2 количественная ПЦР
Для быстрой и надежной оценки репликации вируса мы использовали метод количественной ПЦР в реальном времени (qPCR). Этот анализ потребует меньшей обработки инфекционного SARS-CoV-2 с потенциально высоким титром в BSL3 по сравнению с анализом бляшек VeroE6, поскольку супернатанты могут быть непосредственно помещены в буфер для лизиса, содержащий тиоцианат гуанидиния, который инактивировал бы вирус как минимум на 4 -5 журнал 10 43 .Для измерения уровней SARS-CoV-2 общую РНК экстрагировали из 100 мкл культурального супернатанта с использованием набора EZ.N.A Total RNA Kit I (Omega Bio-Tek) и элюировали 50 мкл воды, свободной от РНКаз. 5 мкл этого экстракта использовали для количественной ПЦР. Были использованы официальные праймеры CDC SARS-CoV-2 N1 и набор зондов TaqMan 44 с набором Luna Universal Probe One-Step RT-qPCR Kit (New England Biolabs):
Прямой праймер: GACCCCAAAATCAGCGAAAT
Обратный праймер: TCTGGTTACTGCCAGTTGAATCTG
TaqMan-зонд: 9TGGTTACTGCCAGTTGAATCTG
Зонда TaqMan: 9T-ACCCCGCATG95CONS-последовательность и последовательность зонда 9T-ACCCCGCATG30AMGTGT5 были 9T-ACCCCGCATG30-AMGTGT9 которые были исследованы.Реакцию кПЦР в реальном времени проводили на термоциклере реального времени Bio-Rad CFX96 в следующих условиях: 55 ° C 10 минут для обратной транскрипции, затем 95 ° C 1 минута с последующими 40 циклами 95 ° C 10 секунд и 60 ° C. C 30-х гг. Абсолютное количественное определение числа копий N1 интерполировали с использованием стандартной кривой с 10 7 -10 1 серийным 10-кратным разведением контрольной плазмиды (nCoV-CDC-Control Plasmid, Eurofins).
Анализ противовирусного ингибирования
Мы использовали ненасыщающую дозу амплифицированного исходного вируса для анализов ингибирования IFN.Ожидалось, что эти титры дадут ~ 10 5 копий на 5 мкл входящего экстракта РНК (дополнительный рис. 1b). Рекомбинантные IFN были получены от PBL Assay Science. В дополнение к IFN-Is (12 подтипов IFNα, IFNβ и IFNω) мы также оценили 3 подтипа IFNλ (IFNλ1, IFNλ2, IFNλ3). Для нормализации IFN мы использовали молярные концентрации 23 вместо международных единиц (МЕ), поскольку значения МЕ были получены из ингибирования вируса энцельфаломиокардита, что может не иметь отношения к SARS-CoV-2.Чтобы найти подходящую дозу для параллельного скрининга 17 IFN, мы провели эксперимент по титрованию дозы штамма USA-WA1 / 2020 с IFNβ и IFNλ1. Доза 2 пМ позволяла максимально различать противовирусную активность IFNβ по сравнению с IFNλ1 (дополнительный рис. 1c). Последовательные 10-кратные разведения IFNβ и IFNλ1 также использовали в последующих экспериментах. Таким образом, в 48-луночных планшетах мы предварительно инкубировали 2,5 × 10 4 клеток A549-ACE2 с IFN в течение 18 часов, затем инфицировали исходным вирусом, амплифицированным A549, в течение 2 часов.После двух промывок PBS добавляли 500 мкл полной среды, содержащей соответствующие IFN. Культуры инкубировали еще 24 часа, после чего супернатанты собирали для выделения РНК и анализа qPCR.
Статистический анализ
Данные анализировали с помощью GraphPad Prism 8. Различия между IFN проверяли с использованием непараметрического двустороннего дисперсионного анализа (ANOVA) с последующим множественным сравнением с использованием критерия Фридмана. Значения коэффициентов корреляции Пирсона (R 2 ) были рассчитаны для анализа линейной регрессии.Парный анализ двух изолятов против нескольких IFN проводили с использованием непараметрического двустороннего рангового критерия согласованных пар Вилкоксона. Различия с p <0,05 считались достоверными.